Capítulo 11: Sistema Endocrino
Definición Regresar
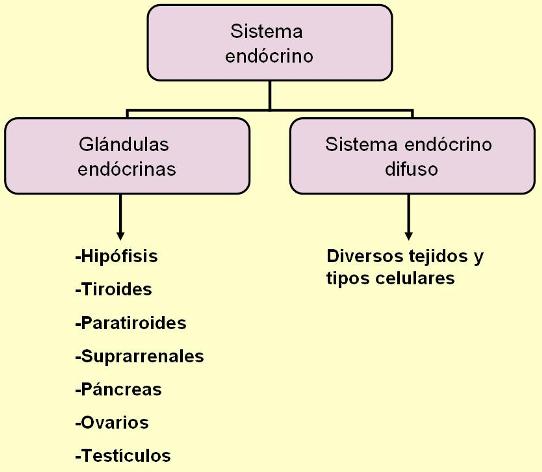
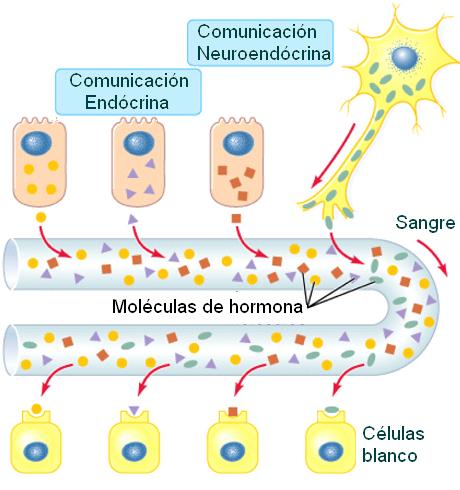
El sistema endócrino es el conjunto de las glándulas endócrinas o glándulas de secreción interna. Las glándulas endócrinas son órganos de origen epitelial especializados en la secreción, que carecen de conductos excretores. Sus secreciones, denominadas hormonas, difunden hacia la sangre y viajan por el torrente circulatorio.
Las hormonas son mensajeros químicos que producen efectos específicos en uno o varios tipos celulares u órganos a los que se llama células u órganos “blanco”. Los blancos de una hormona son aquellas estructuras que poseen un receptor específico para la misma.
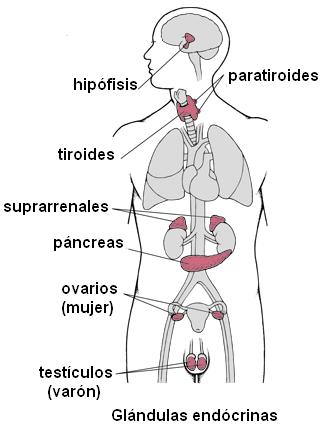
Las glándulas endócrinas son: la hipófisis o pituitaria, la tiroides, las paratiroides, las glándulas adrenales o suprarrenales, una porción del páncreas y las glándulas sexuales (ovarios y testículos).
Además de las glándulas endócrinas propiamente dichas, se incluyen dentro del sistema endócrino diversas células con la capacidad de liberar señales químicas que, ya sea a través del líquido intersticial o de la sangre, llegan hasta otras células blanco donde inducen una respuesta. Podría hablarse entonces de la existencia de un sistema endócrino difuso, formado por todas las células o tejidos corporales que, sin formar parte de una glándula endócrina, secretan mensajeros químicos hacia el medio interno.
Algunos componentes del sistema endócrino difuso son:
- Aparato digestivo: secreta un gran número de señales que controlan las funciones digestivas, como la colecistoquinina o el péptido VIP.
- Corazón: sintetiza el factor natriurético atrial, que controla la presión arterial.
- Riñón: produce eritropoyetina, que estimula la eritropoyesis, y renina, implicada en el control de la presión arterial.
- Endotelio: secreta prostaglandinas y muchas otras señales.
- Leucocitos: producen citoquinas, que modulan la respuesta inmune.
- Tejido adiposo: libera leptina, hormona que interviene en el control del apetito.
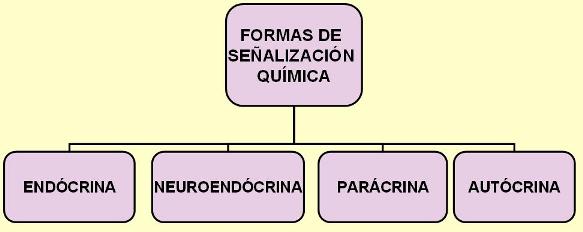
También ciertas neuronas pueden emitir señales químicas que se transportan por sangre. A las hormonas liberadas por células nerviosas se las denomina neurohormonas o secreciones neuroendócrinas.
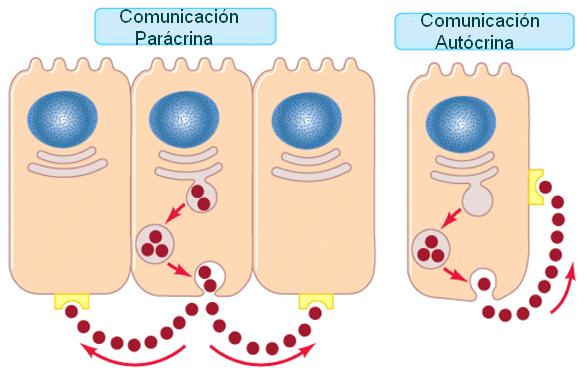
Las hormonas que difunden por el líquido intersticial y ejercen efectos sobre células vecinas reciben el nombre de secreciones parácrinas. En algunos casos, la célula endócrina es el blanco de su propia hormona. A este tipo de señalización se la denomina autócrina.
 |
 |
 |
 |
Glándula hipófisis y eje hipotálamo-hipofisario Regresar
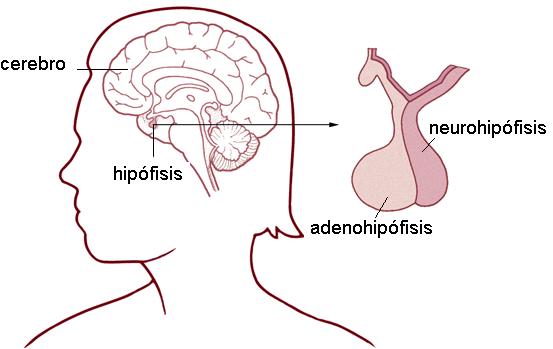
La glándula hipófisis o pituitaria tiene el tamaño de un guisante y se halla ubicada en el cráneo, apoyada sobre la silla turca del hueso esfenoides.
La hipófisis está formada por dos lóbulos:
• Lóbulo anterior o adenohipófisis (adeno: glándula). Éste comprende las porciones tuberal, distal e intermedia.
• Lóbulo posterior o neurohipófisis.
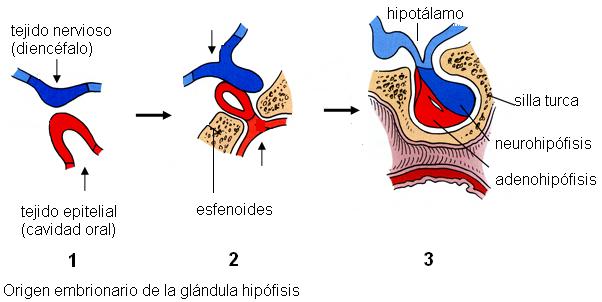
En el embrión, la adenohipófisis se origina a partir del epitelio de la cavidad oral y está formada por tejido glandular, mientras que la neurohipófisis surge como una excrecencia del diencéfalo (una parte del cerebro) y está formada por tejido nervioso. La neurohipófisis permanece conectada al hipotálamo, un núcleo gris del diencéfalo, a través del infundíbulo o tallo pituitario.
Además de la relación ontogénica (por su origen embrionario) y anatómica, el hipotálamo y la hipófisis mantienen una estrecha conexión funcional; las funciones de la hipófisis dependen directamente del hipotálamo y ambos forman una unidad fisiológica: el eje hipotálamo-hipofisario.
La relación funcional entre la hipófisis y el hipotálamo tiene dos aspectos:
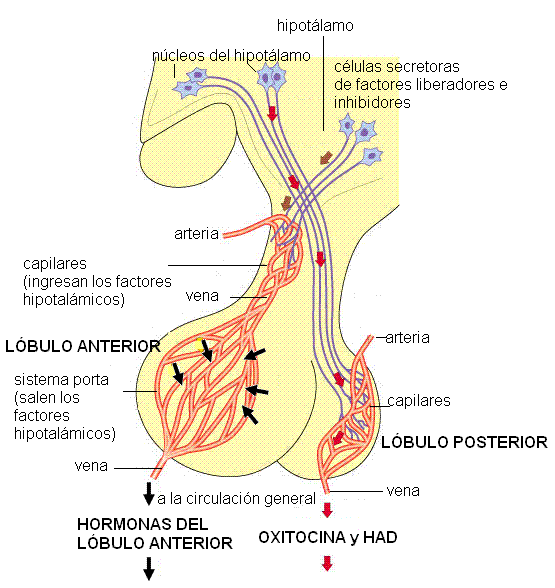 |
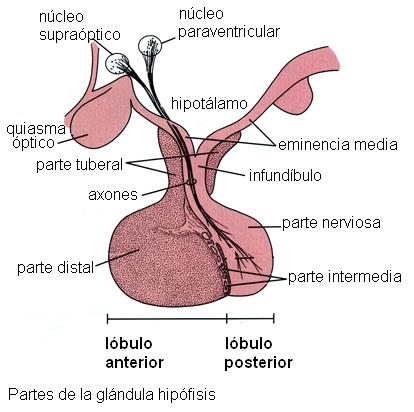
1) La secreción de las hormonas sintetizadas en la adenohipófisis está bajo el control de factores liberadores o inhibidores producidos en el hipotálamo. Estos factores hipotalámicos son sintetizados en neuronas del hipotálamo y llegan hasta las células glandulares del lóbulo anterior mediante la circulación (comunicación neuroendócrina). La comunicación neuroendócrina existente entre el hipotálamo y la adenohipófisis depende del sistema porta hipofisario.
Un sistema porta es una red capilar interpuesta entre dos venas o dos arterias, a diferencia de los capilares comunes, que se interponen entre una arteria y una vena. Los capilares que drenan la sangre del hipotálamo reciben las neurosecreciones y luego se unen formando venas. Estas venas se capilarizan nuevamente al llegar al lóbulo anterior. La segunda red capilar (sistema porta) permite la difusión de los factores hipotalámicos hacia el espacio intersticial y desde allí hacia las células glandulares de la hipófisis anterior, a las cuales controlan. Esta conexión corta y directa entre el hipotálamo y la hipófisis favorece una respuesta rápida, impidiendo que la escasa cantidad de neurohormonas se diluya en la circulación general.
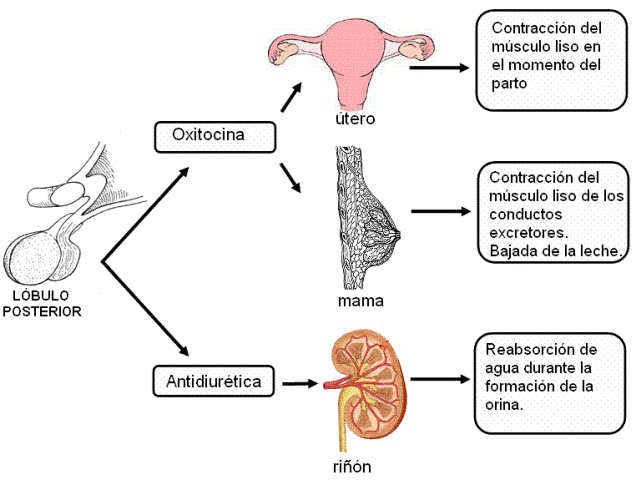
2) La neurohipófisis libera dos hormonas, la oxitocina y la hormona antidiurética (HAD). Estas hormonas son sintetizadas en los cuerpos de neuronas ubicadas en núcleos del hipotálamo (núcleos supraóptico y paraventricular). Una vez sintetizadas descienden por los axones de dichas neuronas, a través del infundíbulo. Los terminales axónicos se encuentran en el lóbulo posterior y desde allí las hormonas son liberadas a la circulación.
Hormonas de la hipófisis Regresar
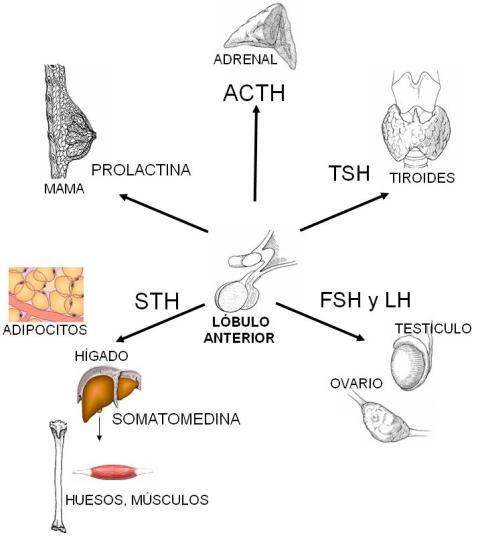
El lóbulo anterior de la hipófisis secreta hormonas tróficas, que actúan estimulando el desarrollo y la función de otros órganos. El lóbulo posterior secreta oxitocina y vasopresina, también llamada hormona antidiurética (HAD). Las hormonas y los efectos específicos sobre sus órganos blanco se resumen en el siguiente cuadro.
|
|
Somatotrofina (STH) |
Es la hormona del crecimiento. Estimula la captación de aminoácidos por las células, promoviendo la síntesis de proteínas.
Estimula la secreción de somatomedinas (factores de crecimiento) en el hígado. Éstas promueven el crecimiento de distintos órganos y especialmente del cartílago, determinando el aumento en longitud de los huesos.
En el tejido adiposo estimula la lipólisis. La liberación de ácidos grasos para la producción de energía produce un ahorro de proteínas, que se destinan al crecimiento.
|
| Prolactina |
Promueve el crecimiento y desarrollo de las glándulas mamarias durante el embarazo. Fomenta la lactancia después del parto.
|
| Adrenocorticotrofina (ACTH) |
Estimula la secreción de glucorticoides y aldosterona en la corteza de la glándula suprarrenal. |
| Tirotrofina (TSH) |
Estimula el crecimiento de la glándula tiroides y la secreción de hormonas tiroideas. |
| Gonadotrofina: Hormona Foliculo-estimulante (FSH) |
Estimula la gametogénesis, tanto en el varón como en la mujer. Regula el ciclo menstrual.
|
| Gonadotrofina: Hormona Luteinizante (LH) |
Estimula la producción de hormonas sexuales, tanto en el varón como en la mujer. Regula el ciclo menstrual. |
 |
 |
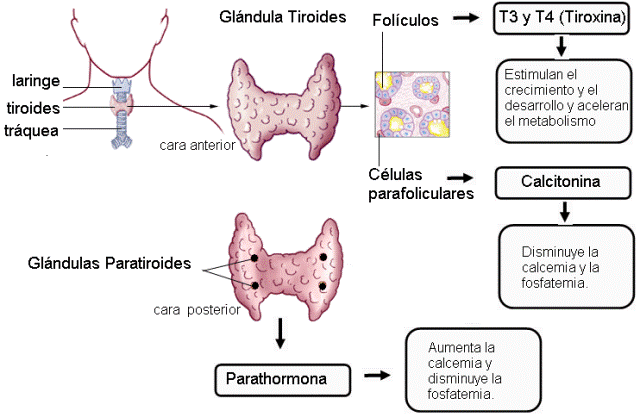
Glándulas tiroides y paratiroides Regresar
La glándula tiroides es bilobulada y está ubicada por delante de la tráquea.
Las glándulas paratiroides son cuatro pequeñas glándulas (dos superiores y dos inferiores) situadas en la cara posterior de la tiroides.
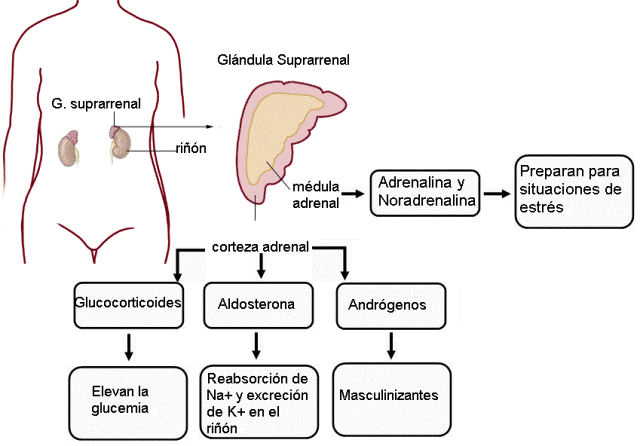
Glándulas suprarrenales Regresar
También llamadas adrenales, son dos glándulas ubicadas en los polos superiores de los riñones. Cada una posee una corteza y una médula, que tienen diferente origen embrionario y diferente función. La corteza está formada por tejido epitelial, en tanto la médula es de origen nervioso. Las secreciones de la médula adrenal son neurohormonas.
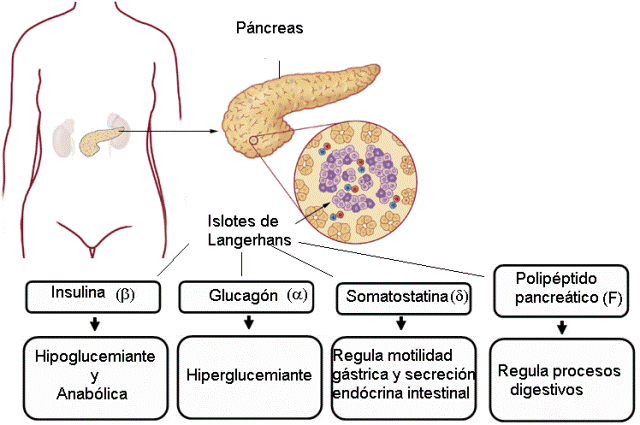
Páncreas Regresar
El páncreas es una glándula mixta. El páncreas exócrino secreta enzimas digestivas que, mediante los conductos pancreáticos, se vuelcan en el intestino delgado.
El páncreas endócrino está conformado por grupos de células endócrinas distribuidos entre el tejido exócrino de la glándula. Las agrupaciones de células endócrinas se conocen como islotes de Langerhans.
En los islotes de Langerhans, distintos tipos celulares (alfa, beta, delta y F) se especializan en la síntesis de diferentes hormonas, como se detalla a continuación.
Regulación del sistema endócrino Regresar
La secreción hormonal es regulada de forma que las concentraciones de hormonas en sangre sean las apropiadas para desencadenar la respuesta necesaria.
Las glándulas endócrinas son reguladas directamente por estímulos nerviosos procedentes de los nervios autónomos.
También son importantes los estímulos bioquímicos, como en el caso de la secreción de insulina, que es disparada por un aumento de la glucemia.
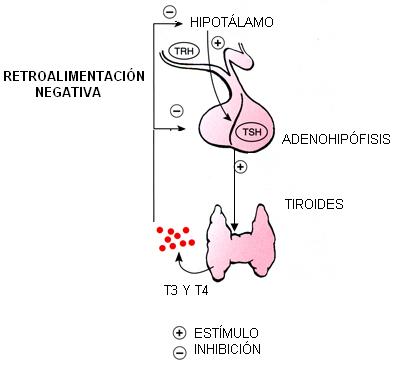
El hipotálamo y la hipófisis regulan la secreción en otras glándulas endócrinas. Tomemos por ejemplo el control en la secreción de hormonas tiroideas.
El hipotálamo secreta un factor liberador (TRH) que estimula a las células de la adenohipófisis especializadas en la síntesis de TSH. La TSH estimula a la glándula tiroides para que sintetice y secrete las hormonas tiroideas, T3 y T4. Pero cuando el nivel en sangre de T3 y T4 es el adecuado, la estimulación por parte del eje hipotálamo-hipofisario debe cesar. De lo contrario, se producirían síntomas por hiperfunción de la glándula.
La hipófisis y, en menor medida, el hipotálamo, detectan el nivel elevado de T3 y T4 en sangre. Ante esta señal, el hipotálamo disminuye la síntesis del factor liberador y la hipófisis frena la secreción de TSH. Así se logra mantener una concentración adecuada de T3 y T4. Este mecanismo de control recibe el nombre de retroalimentación negativa: el efecto (nivel de T3 y T4) suprime la causa (factor liberador y TSH).
Cuando los niveles de T3 y T4 descienden más allá de lo conveniente, el hipotálamo y la hipófisis reanudan sus secreciones y el ciclo se reinicia.
 |
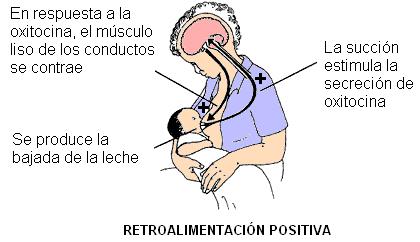
Un tipo de regulación diferente se produce en el caso de la oxitocina. Durante el embarazo, la prolactina y otras hormonas estimulan el desarrollo de la mama, preparándola para la producción de leche. Sin embargo, la leche no baja hasta que se coloca el bebé al pecho y éste comienza a succionar. La eyección de la leche depende de la contracción del músculo liso de la glándula, estimulado por la oxitocina. La secreción de oxitocina es la respuesta de un reflejo nervioso disparado por el estímulo de la succión.
En este ejemplo se observa la integración de los controles nervioso y endócrino, y además un tipo de retroalimentación positiva.
Cuanto más se estimula el pezón, mayor es la secreción de oxitocina y mayor la cantidad de leche secretada. Por eso, las mujeres pueden amamantar por largos períodos, mientras mantengan la frecuencia de la lactancia. Si, en cambio, dejan de amamantar o disminuyen la frecuencia, el estímulo es cada vez menor y la secreción de leche empieza a disminuir hasta que cesa.
 |
Composición química de las hormonas Regresar
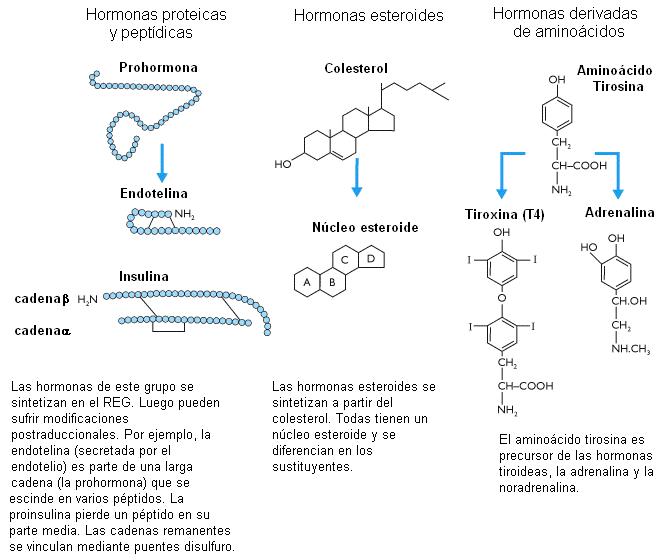
Desde el punto de vista químico, las hormonas pueden agruparse en cuatro grandes clases:
1) Péptidos, proteínas, glucoproteínas: pueden ser cadenas muy cortas o de alto PM. Se sintetizan en el sistema de endomembranas y se almacenan en gránulos secretorios hasta la exocitosis. Incluyen a las hormonas de hipotálamo, hipófisis, páncreas y paratiroides.
2) Derivados de aminoácidos: son pequeñas moléculas hidrosolubles. Se sintetizan en el citosol y luego se introducen en vesículas donde son reservadas. Por ejemplo: hormonas tiroideas.
3) Esteroides: derivan del colesterol. Son liposolubles. Se sintetizan en REL y mitocondrias. No se almacenan. Son las hormonas corticoadrenales y sexuales y la vitamina D.
4) Eicosanoides: derivan de un ácido graso (ácido araquidónico). Se sintetizan en la membrana plasmática y no se almacenan. Actúan como mensajeros locales (parácrinos). Son las prostaglandinas y los leucotrienos, producidos en casi todos los tipos celulares.
Mecanismo de acción de las hormonas en las células blanco Regresar
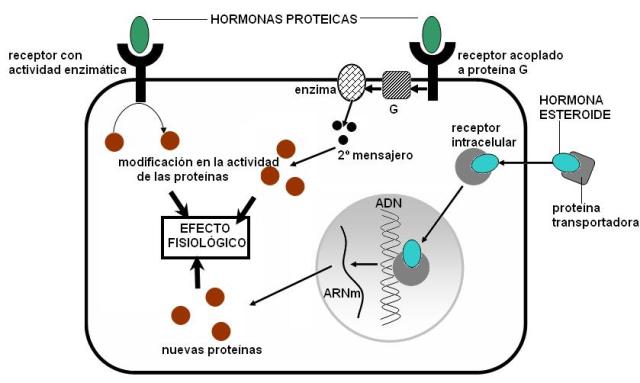
Las hormonas ejercen sus efectos a través de su unión al receptor. Los receptores de hormonas son proteínas. Cuando se unen la hormona y el receptor, este último experimenta un cambio conformacional. El receptor activado por la hormona inicia la vía de “transducción de la señal”. Ésta consiste en una cascada de modificaciones en la fisiología de la célula blanco que, directa o indirectamente, produce lo que llamamos la “respuesta” a la hormona.
Cada tipo de receptor dispara una vía de transducción particular. A su vez, esta vía es diferente según el tipo de célula. Dado que para una hormona pueden existir varios receptores distintos, esto explica la diversidad de respuestas inducidas por la misma hormona en los órganos blanco.
Hay dos grandes tipos de receptores hormonales: los que se encuentran insertos en la membrana plasmática y los intracelulares.
Los receptores de membrana reconocen a hormonas de naturaleza hidrofílica, por ejemplo a las hormonas proteicas. Este tipo de moléculas, poco afines a los lípidos, no puede atravesar la membrana celular y por lo tanto el receptor debe estar ubicado en la superficie de la célula blanco.
Las hormonas lipofílicas, como los esteroides, difunden fácilmente a través de las bicapas lipídicas. Estas hormonas traspasan la membrana plasmática y se unen a un receptor intracelular.
Una vez formado el complejo hormona-receptor se desencadena el efecto fisiológico en la célula blanco.
a) En algunos casos, los receptores de membrana tienen una actividad enzimática que es “encendida” por la llegada de la hormona. Estos receptores catalizan la fosforilación (adición de grupos fosfato) de proteínas celulares, que a su vez se activan o se inactivan (según el caso) al ser fosforiladas. De esta forma se modifica la actividad celular.
b) Otros receptores de membrana actúan acoplados a una proteína G. Las proteínas G (de las que existen varios tipos) son las encargadas de inhibir o activar enzimas que fabrican “segundos mensajeros”. Los segundos mensajeros son como los “voceros” de la hormona (el primer mensajero) en el medio intracelular. La respuesta de la célula blanco depende del control que los segundos mensajeros ejercen sobre ciertas proteínas celulares. Un nucleótido derivado del ATP, el AMP cíclico (AMPc) y el calcio actúan frecuentemente como segundos mensajeros.
c) El receptor de hormonas esteroides se une a la hormona en el citoplasma. La unión de la hormona con el receptor capacita al complejo para ingresar al núcleo. Dentro del núcleo, el receptor interactúa con regiones reguladoras del ADN, estimulando la transcripción de genes específicos. Las nuevas proteínas sintetizadas a partir de dichos genes son las causantes del efecto fisiológico en la célula blanco.
La respuesta celular a las hormonas proteicas o peptídicas es mucho más rápida que la respuesta a hormonas esteroideas. Esto se debe a que las primeras actúan a través de la activación de proteínas preexistentes, en cambio las hormonas esteroideas inducen la síntesis de nuevas proteínas. Insume más tiempo fabricar una proteína desde el inicio que simplemente activarla.
Comparación de los sistemas endócrino y nervioso Regresar
El sistema nervioso y el endócrino trabajan estrechamente ligados. Existen muchos mecanismos por los cuales uno afecta la función del otro y en algunos casos sus funciones se solapan.
Los dos sistemas tienen características comunes. Ambos actúan por intermedio de mensajeros químicos, los neurotransmisores y las hormonas, aunque éstos viajan de manera diferente.
Tanto los neurotransmisores como las hormonas inducen efectos en células diana donde se localizan receptores específicos.
Las respuestas endócrinas son relativamente lentas, pero prolongadas. Las respuestas nerviosas son muy rápidas y también más fugaces.
El sistema nervioso permite respuestas a estímulos externos e internos y gobierna la relación con el ambiente. El sistema endócrino también está sometido a la influencia del ambiente, pero de manera indirecta, por intermedio del sistema nervioso. El sistema endócrino ejerce fundamentalmente el control del medio interno. Regula el metabolismo, la presión arterial, el crecimiento y el desarrollo, la reproducción y ciertos aspectos de la conducta.
|