|
Capítulo 8: Sistema Inmunitario
Introducción Regresar
Inmunidad significa “defensa”. El sistema inmunitario comprende a numerosas estructuras, órganos y células, que se distribuyen en todo el cuerpo y nos protegen contra las agresiones de los patógenos: virus, bacterias, hongos y otros microorganismos con los que estamos en continuo contacto y que nos pueden causar enfermedad.
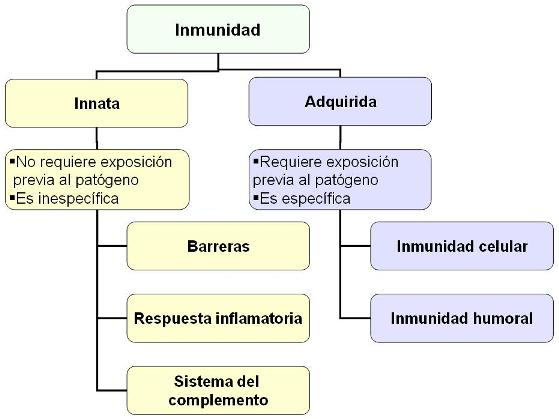
Las defensas del organismo se pueden clasificar en dos tipos: inmunidad innata e inmunidad adquirida.
La inmunidad innata es constitutiva. Abarca una serie de estructuras y mecanismos que forman parte de la constitución del individuo. Los mecanismos de defensa innatos son inespecíficos, nos defienden de cualquier invasor y no dependen de la exposición previa del individuo a los patógenos. Proporcionan inmunidad innata las barreras mecánicas y químicas, la respuesta inflamatoria y el sistema del complemento.
Los mecanismos de la inmunidad adquirida, en cambio, se ponen en marcha durante la primera exposición a un patógeno y protegen frente a futuros encuentros con el mismo tipo de patógeno. La inmunidad adquirida es específica.
A pesar de estas diferencias, el sistema inmunitario es uno solo y trabaja en forma integrada. Los mecanismos innatos y los adquiridos actúan simultáneamente, y muchos componentes del sistema inmune participan en ambos tipos de respuesta.
 |
Barreras mecánicas y químicas Regresar
| |
Barreras Mecánicas |
Barreras Químicas |
| |
|
- Lisozima
- Sebo
- Sudor
- pH
- Lactoferrina
|
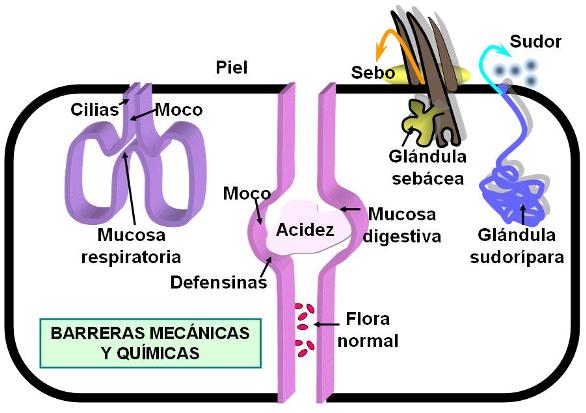
Las barreras mecánicas son aquellas estructuras que evitan la entrada de los patógenos.
La primera barrera defensiva del cuerpo es la piel. El epitelio pluriestratificado queratinizado que forma la epidermis, cuando está íntegro, actúa como una eficaz protección que impide el ingreso de los gérmenes.
Los epitelios que tapizan la luz de los sistemas conectados al exterior, como el digestivo, el respiratorio, el reproductor y el urinario, también presentan adaptaciones destinadas a la defensa.
Estos epitelios son “mucosas”, capas celulares que contienen células secretoras de moco, el cual forma un tapiz sobre la superficie apical del epitelio. La capa de moco dificulta la llegada de los patógenos hasta la superficie celular.
La capa de moco también contiene defensinas. Las defensinas son péptidos que destruyen bacterias, hongos y parásitos.
El epitelio respiratorio posee además células ciliadas. El batido coordinado de las cilias ayuda a “barrer” el moco donde quedan retenidas las partículas que ingresan a la vía respiratoria junto con el aire.
Las mucosas, de todos modos, no son tan inexpugnables como la piel y suelen ser las puertas de entrada más frecuentes para los agentes infecciosos o sus toxinas.
Además de las barreras mecánicas, la piel y las mucosas, el cuerpo cuenta con barreras químicas. Muchas sustancias presentes en las secreciones corporales tienen actividad antimicrobiana. Por ejemplo, la lisozima, una enzima que se encuentra en las lágrimas y en la saliva, degrada las paredes celulares de algunas bacterias. En el sebo y en el sudor hay otras sustancias antibióticas, que impiden el desarrollo de los microbios.
El pH de las secreciones puede inhibir el desarrollo de patógenos. Las secreciones vaginales tienen pH ácido. El ácido clorhídrico secretado por la mucosa gástrica (del estómago), acidifica el jugo gástrico, determinando un pH =2, que mata a la casi totalidad de los microorganismos presentes en los alimentos.
En las personas desnutridas, ante la escasez de alimentos, el organismo reacciona con un “ajuste” tendiente a economizar los recursos. Algunas funciones se ven disminuidas para salvaguardar aquéllas que son esenciales para la supervivencia. Entre las funciones afectadas está la secreción de jugo gástrico. Ésta es una de las razones por las cuales las personas desnutridas resultan más susceptibles a enfermedades de transmisión alimentaria, como el cólera.
En la leche y otras secreciones se encuentra la lactoferrina. Esta proteína se une específicamente al hierro, disminuyendo la disponibilidad del mismo para las bacterias que lo utilizan como nutriente.
Por último, la existencia de una flora normal, formada por bacterias que se alojan en nuestro cuerpo en una convivencia pacífica, es otro elemento protector. Estas bacterias, que usualmente no nos perjudican, excluyen, por competencia, a otras que son decididamente patógenas.
 |
Respuesta inflamatoria Regresar
Cuando las barreras defensivas fallan y los tejidos son invadidos por agentes patógenos, se activa la segunda línea de defensa: la respuesta inflamatoria.
La respuesta inflamatoria es un espectacular cambio en el tejido dañado, provocado por muchos mediadores químicos, tanto del propio tejido como de los gérmenes invasores.
Una sustancia liberada durante la inflamación es la histamina, secretada por los leucocitos basófilos y las células cebadas del tejido conectivo. La histamina tiene un efecto vasodilatador, lo que ocasiona que la zona dañada reciba un mayor flujo sanguíneo. Simultáneamente, aumenta la permeabilidad capilar y se produce un escape de líquido desde la sangre hacia el espacio intersticial. Los síntomas y signos clásicos de la inflamación, conocidos como la tétrada de Celsius (el médico que los describió) son “rubor, calor, tumor (hinchazón) y dolor”. Todos ellos son causados por la mayor llegada de sangre y el aumento de la permeabilidad en la zona afectada.
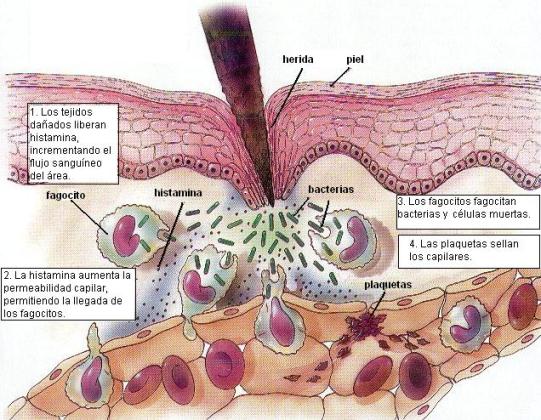 |
Con el mayor riego sanguíneo llegan también gran cantidad de neutrófilos y monocitos, llamados en conjunto “fagocitos”. Los fagocitos poseen cuatro propiedades que son fundamentales en la respuesta inflamatoria: quimiotaxis, diapédesis, ameboidismo y fagocitosis. La quimiotaxis es el fenómeno de atracción que ciertas sustancias ejercen sobre las células, haciendo que éstas se movilicen hacia la fuente de las mismas, donde su concentración es mayor. Los fagocitos son atraídos hacia el tejido por los mediadores liberados y también por toxinas bacterianas.
La diapédesis es la capacidad de atravesar las paredes capilares. Una vez en la zona de inflamación, los fagocitos se desplazan mediante movimientos ameboideos. Los movimientos ameboideos (por ejemplo la ameba, un protozoo que se traslada de esta forma) consisten en el desplazamiento de la célula sobre un sustrato, mediante la emisión de prolongaciones citoplasmáticas llamadas pseudópodos. En el movimiento ameboideo tienen participación los filamentos de actina, que dirigen la formación de pseudópodos, y las uniones de adhesión ente las células y la matriz extracelular.
La función más importante de los fagocitos es la fagocitosis, que significa “ingestión celular”. La fagocitosis es un tipo de transporte en masa que permite la incorporación de grandes partículas sólidas, las cuales son rodeadas por los pseudópodos hasta que quedan completamente englobadas en una vesícula (el fagosoma). Por medio de la fagocitosis, neutrófilos y macrófagos “comen” selectivamente a los agentes invasores y también a los restos celulares del tejido dañado.
La selectividad en la fagocitosis se relaciona con la expresión, en la membrana plasmática de los fagocitos, de receptores específicos que reconocen algunas moléculas comunes a muchos patógenos (Receptores de Reconocimiento de Patrón asociados a patógenos = RRP).
Los fagosomas se fusionan posteriormente con los lisosomas y de esta forma se digiere su contenido, gracias a las hidrolasas lisosomales. También contribuye la mieloperoxidasa lisosómica que sintetiza hipoclorito, un potente bactericida. Además, las membranas de los fagosomas ensamblan sistemas enzimáticos que forman agentes oxidantes, como el peróxido de hidrógeno, mortales para las bacterias.
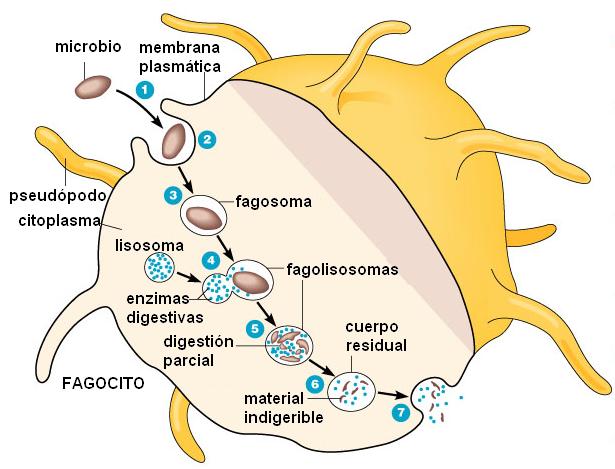 |
- Quimiotaxis y adherencia del microbio al fagocito.
- Ingestión del microbio por el fagocito.
- formación del fagosoma.
- Fusión del lisosoma primario con el fagosoma. Formación del fagolisosoma (lisosoma secundario).
- Digestión del microbio.
- Formación del cuerpo residual con el material indigerible.
- Exocitosis del contenido del cuerpo residual.
|
Cuando el patógeno no puede ser atacado por una sola célula, es rodeado por grupos de leucocitos que exocitan defensinas y todo su arsenal de enzimas y sustancias tóxicas.
Algunas bacterias tienen paredes o cápsulas celulares que resisten la fagocitosis y otras se defienden con sustancias que destruyen la membrana lisosómica.
Los neutrófilos se desactivan después de fagocitar unas 20 bacterias y los macrófagos pueden fagocitar hasta 100. La mayoría de los neutrófilos y algunos macrófagos finalmente mueren y se acumulan junto con el tejido necrótico y el líquido extracelular. Esta mezcla forma el pus. La mieloperoxidasa, que contiene cobre en su estructura, es la responsable del color verdoso del pus.
Las zonas inflamadas se rodean con coágulos de fibrinógeno, el cual se extravasa al aumentar la permeabilidad capilar. Los coágulos tabican la zona y taponan los capilares, inhibiendo así el desarrollo de los microbios (que quedan privados de nutrientes) y retrasando su diseminación. La coagulación es inducida por citoquinas, hormonas secretadas por los leucocitos.
Las citoquinas también inducen una respuesta sistémica, que acompaña a todos los fenómenos locales descritos. Por ejemplo, algunas citoquinas actúan sobre el hipotálamo, donde se localiza el centro regulador de la temperatura, y causan fiebre. La fiebre es un mecanismo defensivo, ya que las altas temperaturas favorecen la respuesta del sistema inmune y en cambio perjudican a los patógenos, que se desarrollan mejor a temperaturas bajas.
La respuesta sistémica implica asimismo un aumento en la síntesis de hormonas esteroides y de las proteínas de fase aguda, que se fabrican en el hígado.
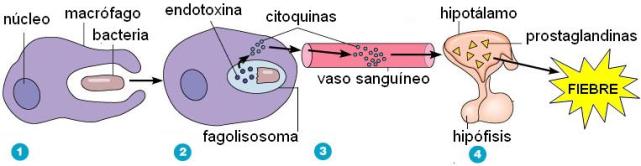 |
| 1) El macrófago fagocita una bacteria. |
2) La bacteria es digerida.Partes de la bacteria (endotoxinas) disparan la secreción de citoquinas.
|
3) Las citoquinas llegan al hipotálamo por la circulaciñon sistémica. |
4) En respuesta a las citoquinas, el hipotálamo secreta prostaglandinas, que producen fiebre. |
| Dos listas clave para caracterizar la respuesta inflamatoria |
| Tétrada de Celsius |
Propiedades de los fagocitos |
|
- Quimiotaxis
- Diapédesis
- Ameboidismo
- Fagocitosis
|
Sistema del complemento Regresar
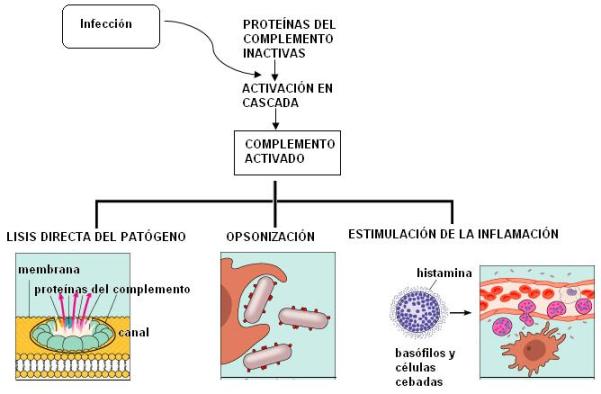
El sistema del complemento es un conjunto de unas 20 proteínas sintetizadas en el hígado, que circulan por la sangre y el líquido extracelular. La mayoría de ellas se encuentran inactivas hasta que se produce una infección.
Las proteínas del complemento se activan en cascada. La activación se produce por proteólisis, es decir por la escisión de la cadena proteica. Cuando una proteína del complemento se escinde, se activa y provoca la escisión de la siguiente.
Una vez activadas, las proteínas del complemento pueden actuar provocando la lisis directa del patógeno. Se introducen en la membrana del patógeno y forman canales que alteran la permeabilidad, causándole la muerte.
También complementan la respuesta inmune específica, mediante un mecanismo denominado opsonización. La opsonización consiste en la adhesión del complemento a la superficie del patógeno, lo que favorece la fagocitosis a cargo de los macrófagos (se suele decir que el complemento es como un condimento que hace más sabroso al patógeno).
Además participan en la respuesta inflamatoria reclutando fagocitos y estimulando la secreción de histamina.
 |
Inmunidad adquirida Regresar
 |
Después de padecer una enfermedad infecciosa, se desarrolla memoria inmunitaria. |
La inmunidad adquirida es un tipo de protección específica contra el patógeno que la estimula, y es duradera. Es el tipo de inmunidad que hace que una persona que padeció una cierta enfermedad infecciosa en la infancia, por ejemplo: sarampión, quede protegida frente al agente patógeno del sarampión para el resto de su vida o al menos por muchos años.
Para poner en marcha los mecanismos de la inmunidad adquirida es fundamental que el organismo reconozca a sus propios componentes como tales y los diferencie de los extraños. |
De hecho, cuando no se diferencia adecuadamente lo propio de lo ajeno, los mecanismos de defensa se vuelven contra el organismo dando origen a las enfermedades autoinmunes.
De la misma forma, también son atacados los transplantes que provienen de otro individuo.
Las células que sufren una transformación maligna (células cancerosas) expresan moléculas que no expresan las células normales. Las células cancerosas son reconocidas y eliminadas por el sistema inmune. Los cánceres que prosperan son los que logran escapar a este sistema de vigilancia.
En algunos casos, el sistema inmune monta una defensa contra partículas extrañas que no son patógenas. A este fenómeno se lo denomina hipersensibilidad y es la base de los procesos alérgicos. Los individuos alérgicos padecen síntomas que obedecen a una respuesta inmune exagerada contra elementos que en realidad son inocuos. El sistema inmune de la mayoría de los individuos ignora a estos “alergenos” (polen, polvo, ácaros, etc.).
| En conclusión, la respuesta inmune adquirida participa en: |
- defensa específica y duradera contra patógenos
- enfermedades autoinmunes
- rechazo de transplantes
- vigilancia inmunológica contra células malignas
- alergias
|
 |
 |
Las enfermedades autoinmunes, como la Diabetes 1, y las alergias, son efectos negativos de la respuesta inmune. |
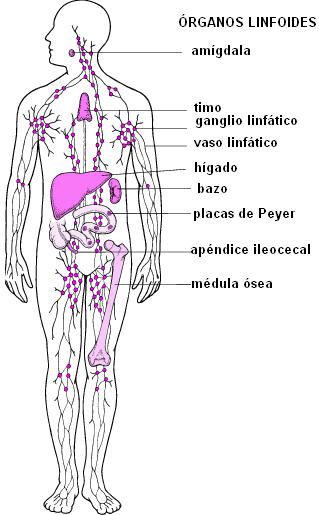
Las células protagonistas de la respuesta inmune adquirida son los linfocitos. originados en los órganos linfoides.
Órganos linfoides primarios y secundarios Regresar
Existen dos clases principales de linfocitos: los linfocitos B (LB) y los linfocitos T (LT). Los linfocitos, como las demás células sanguíneas, se originan a partir de células pluripotenciales, en los órganos linfoides primarios.
Para los linfocitos B, los órganos linfoides primarios son los órganos hematopoyéticos: el hígado fetal y la médula ósea en el adulto. (En las aves, los LB se diferencian en un órgano llamado bolsa de Fabricio, de donde surge la “B” de la denominación).
Los linfocitos T se originan en el timo, hacia donde migran las células pluripotenciales desde los órganos hematopoyéticos. El timo es una glándula ubicada en la región anterior del tórax.
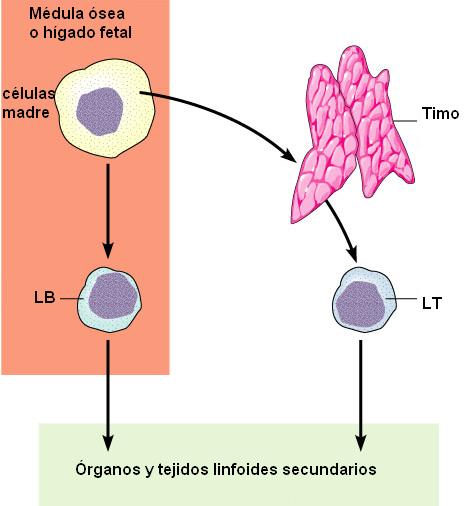 |
Una vez formados, los linfocitos circulan en la sangre y en la linfa y se acumulan en otros órganos, llamados órganos linfoides secundarios, donde se produce el encuentro con los patógenos. Los órganos linfoides secundarios son el bazo, los ganglios linfáticos, el apéndice ileocecal, las amígdalas y los nódulos, acumulaciones de linfocitos en las paredes de muchos otros órganos (como las placas de Peyer del intestino).
Complejo mayor de histocompatibilidad Regresar
El complejo mayor de histocompatibilidad (CMH) es un conjunto de proteínas de membrana que cobra una gran importancia en los mecanismos de inmunidad adquirida.
Hay dos tipos de CMH: CMH de clase I y CMH de clase II.
Prácticamente todas las células nucleadas del cuerpo expresan el CMH de clase I. El CMH de clase II se expresa solamente en algunas células del sistema inmunitario.
 |
 |
| El CMH funciona como un carnet de identidad para las células propias. |
Las proteínas que forman el CMH están codificadas por numerosos genes. Un individuo hereda una combinación particular de estos genes, por lo cual el CMH es diferente en cada individuo de una misma especie. Antígeno Regresar
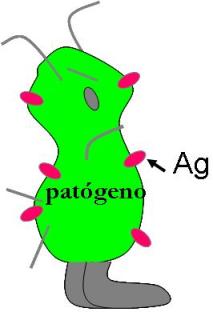
Se denomina antígeno (Ag) a moléculas que el organismo reconoce como extrañas y que, por lo tanto, estimulan la respuesta inmune. Los antígenos son, generalmente, partes de los gérmenes patógenos.
Linfocitos T e inmunidad celular Regresar
Los linfocitos T son los efectores de un tipo de inmunidad adquirida denominada inmunidad celular o inmunidad mediada por células.
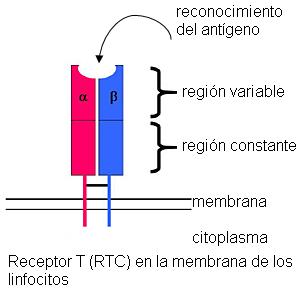
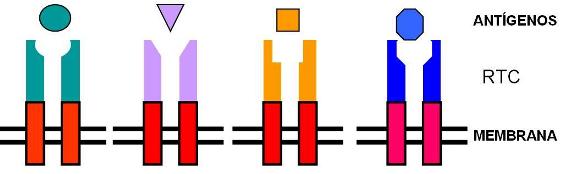
Los linfocitos T poseen en sus membranas un receptor, el TCR (receptor de células T). Una parte de este receptor es siempre igual (región constante). Otra parte del receptor es la región variable. La región variable presenta una enorme diversidad en distintas poblaciones de linfocitos T. Esta región es la encargada de reconocer al antígeno.
La diversidad de TCR es tan grande, que existen en el organismo poblaciones de linfocitos T para reconocer específicamente a millones de Ag distintos.
Cuando las diferentes poblaciones de LT se diferencian en el timo, se eliminan aquellas cuyos receptores podrían reconocer a moléculas propias, evitando así que la respuesta inmune celular se ejerza contra el mismo organismo (autotolerancia).
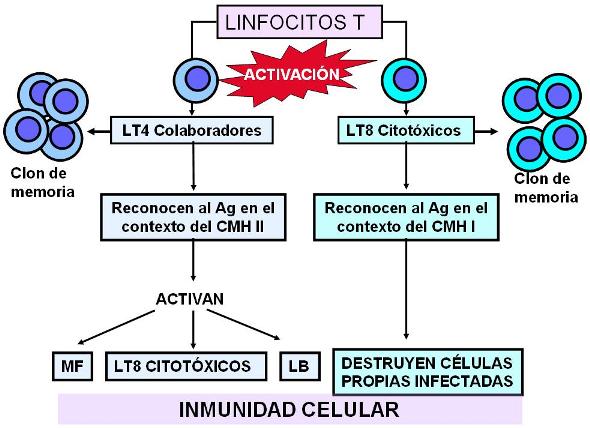
Existen dos clases de linfocitos T: los colaboradores (“helpers”) y los citotóxicos.
Para que un linfocito T colaborador inicie la respuesta inmune celular debe ser activado por una “célula presentadora de antígeno” (CPA). Las CPA son los macrófagos y otras células emparentadas, llamadas células dendríticas.
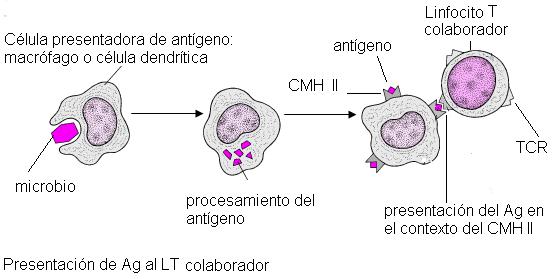 |
Cuando las CPA fagocitan un germen, lo digieren, y luego exhiben en sus membranas moléculas peptídicas procedentes de esos gérmenes, que se comportan como antígenos.
Los antígenos son presentados en la membrana de las CPA unidos a moléculas del CMH II, complejo mayor de histocompatibilidad de clase II.
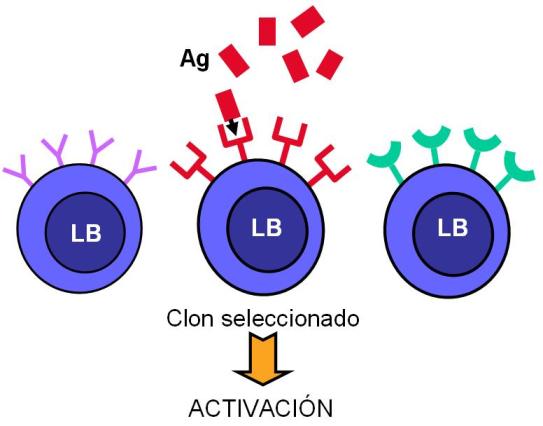
En los órganos linfoides secundarios, las CPA interactúan con los linfocitos T colaboradores y les presentan los antígenos en el contexto del CMH de clase II. De esta forma los LT son activados y se convierten en células efectoras. Las CPA no activan a todos los LT; el LT activado es aquél cuyo TCR tiene la estructura adecuada para reconocer específicamente al antígeno que le está siendo presentado. A éste se lo llama linfocito “competente”. La activación selectiva de la población de linfocitos competentes entre todas las poblaciones existentes se conoce como “selección clonal”.
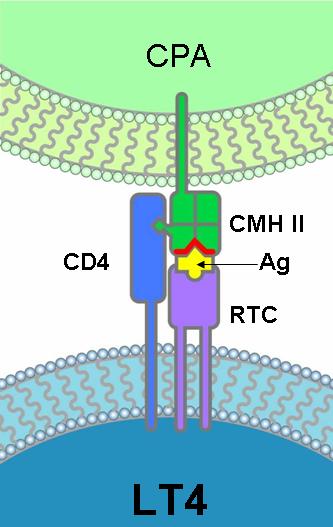
Junto al TCR los linfocitos T colaboradores presentan un correceptor, el CD4, que reconoce al CMH de clase II.
 |
| Los linfocitos T colaboradores activados se convierten en efectores con diversas funciones: |
• Activan a los macrófagos para que resulten más efectivos en la destrucción de microorganismos intracelulares.
• Colaboran con los linfocitos T citotóxicos.
• Colaboran con los linfocitos B.
|
La activación puede producirse por contacto célula-célula (mediante receptores de membrana específicos) o por medio de la secreción de citoquinas.
Si bien los LT colaboradores no destruyen patógenos en forma directa, resultan esenciales para llevar a cabo la respuesta inmune, por su papel activador de otras células.
|
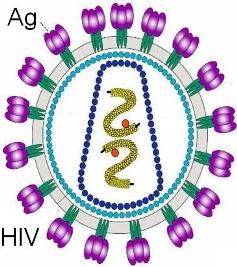
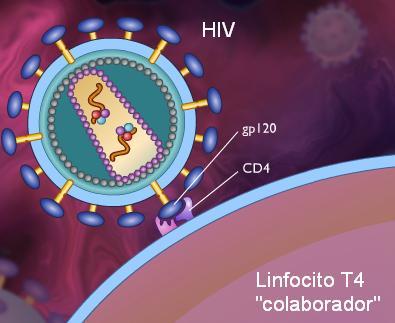
El virus que causa el SIDA, el HIV, utiliza un receptor celular para ingresar a sus células huésped: el CD4. Por lo tanto, los linfocitos colaboradores son el blanco predilecto del HIV. Al reproducirse, el HIV destruye a los LT colaboradores, desbaratando la respuesta inmune. SIDA significa “Síndrome de Inmunodeficiencia Adquirida”. Las personas infectadas por HIV son propensas a padecer todo tipo de infecciones por la pérdida de sus LT colaboradores. |
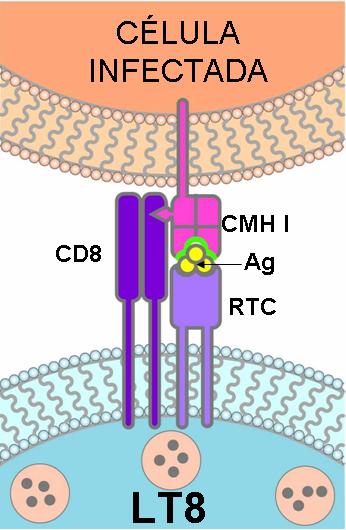
Otro tipo de linfocito T son los citotóxicos. Los linfocitos T citotóxicos activados se especializan en la destrucción de células propias infectadas por parásitos intracelulares.
Los virus y algunas bacterias son parásitos intracelulares. Cuando una célula alberga a este tipo de parásitos, moléculas antigénicas procedentes de los mismos son procesadas y presentadas en el CMH de clase I de dicha célula. De esta forma, la célula infectada es reconocida por los linfocitos T citotóxicos.
Los linfocitos T citotóxicos efectúan un doble reconocimiento:
- reconocen al antígeno a través del receptor de célula T (TCR), que es específico para ese antígeno, y
- reconocen a la célula propia por la presencia del CMH de clase I. Para el reconocimiento del CMH de clase I los linfocitos T citotóxicos poseen un correceptor denominado CD8.
 |
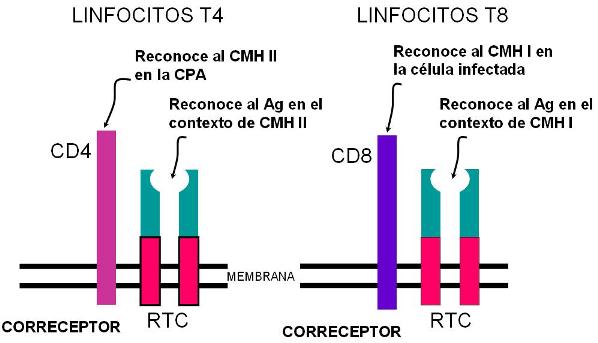 |
DOBLE RECONOCIMIENTO: tanto los linfocitos T4 como los T8 efectúan un doble reconocimiento. El RCT interacciona con el Ag, mientras que el correceptor interacciona con una molécula del MHC. |
| Acción de los LT citotóxicos |
Una vez efectuado el reconocimiento, los linfocitos T citotóxicos destruyen a la célula infectada. Para hacerlo, utilizan distintas estrategias que terminan induciendo a la célula a la apoptosis.
La apoptosis es una especie de suicidio celular. La maquinaria para la apoptosis consta de proteínas llamadas caspasas, que normalmente están inactivas. El contacto del linfocito T citotóxico con la célula infectada activa una cascada de caspasas que conducen a la muerte celular.
Una de las estrategias utilizadas por los linfocitos para activar la cascada de caspasas consiste en la secreción de perforinas. Las perforinas se introducen en la membrana plasmática de la célula blanco y forman un canal (semejante al que produce el sistema del complemento). A través del canal ingresan otros componentes que activan las caspasas. |
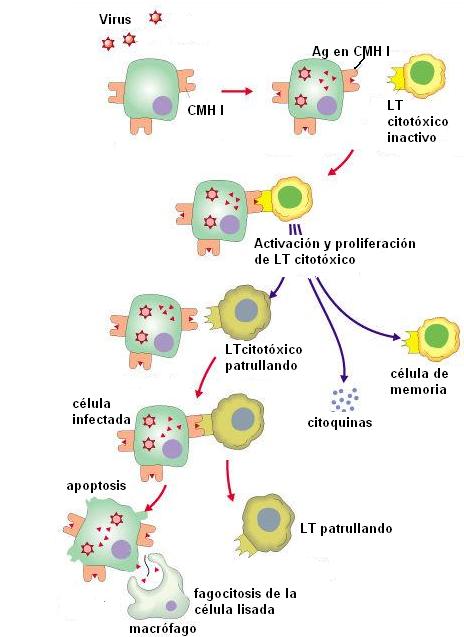 |
Una característica de la respuesta inmune, tanto celular como humoral, es la memoria. Esto se debe a que los linfocitos activados, además de convertirse en efectores, proliferan formando clones de células de memoria. Éstas perduran, posibilitando una defensa más rápida en posteriores encuentros con el mismo tipo de antígeno que activó la respuesta.
Linfocitos B e inmunidad humoral Regresar
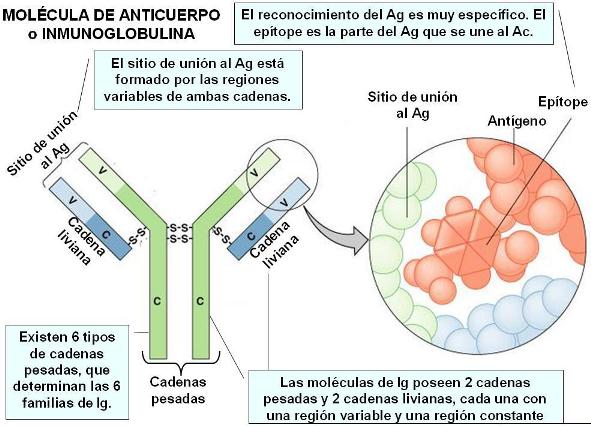
Los linfocitos B, homólogamente a los linfocitos T, presentan receptores específicos para la detección de antígenos. Los receptores de los linfocitos B son proteínas denominadas anticuerpos (Ac) o inmunoglobulinas (Ig).
Las inmunoglobulinas tienen una región constante (C) y una región variable (V). Diferencias en la región constante determinan la existencia de 5 familias de Ig: Ig A, Ig D, Ig G, Ig E e Ig M.
La región variable exhibe una enorme diversidad, que permite el reconocimiento específico de millones de antígenos diferentes.
Los linfocitos B son activados en los órganos linfoides secundarios por los LT colaboradores o bien por contacto con el antígeno, mediante un proceso de selección clonal.
La activación del LB competente induce su proliferación. Algunas de las células hijas permanecen como células de memoria, mientras que otras se diferencian en células plasmáticas o plasmocitos. Las células plasmáticas diferenciadas a partir del LB activado secretan inmunoglobulinas que ingresan a la circulación.
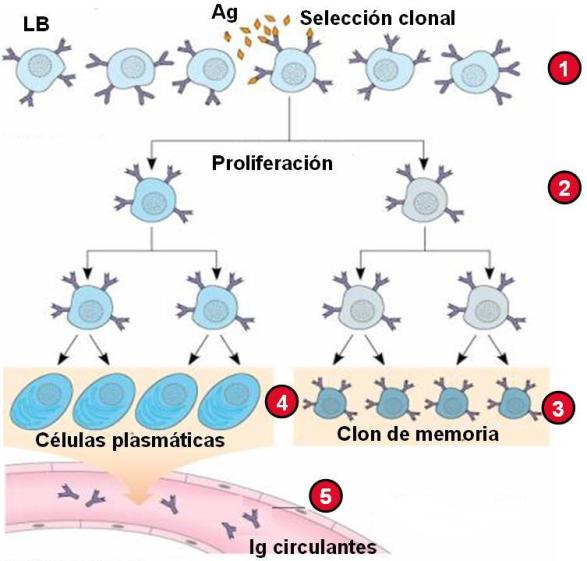 |
1) El LB competente se une al Ag (selección clonal). 2) El LB se activa y prolifera. 3) Algunos LB se diferencian a células de memoria. 4) Otros LB se diferencian a células plasmáticas. 5) Las células plasmáticas secretan Ig (anticuerpos) a la circulación. |
Las células plasmáticas son células especializadas en la síntesis de inmunoglobulinas con la misma especificidad antigénica que la célula madre, es decir, capacitadas para reconocer al mismo tipo de antígeno que activó la respuesta. Sin embargo, las inmunoglobulinas ya no permanecen ancladas a la membrana del linfocito, sino que son secretadas. Este cambio en el tipo de inmunoglobulina producida (de molécula de membrana a molécula de secreción) se debe a un diferente procesamiento del ARNm de las inmunoglobulinas. El mismo ARNm puede ser poliadenilado en dos puntos diferentes, determinando un ARNm maduro largo o un ARNm maduro corto. El primero es traducido en una proteína con una región hidrofóbica carboxiterminal, que sirve para insertar a la inmunoglobulina en la membrana. Cuando se traduce el ARNm más corto, la cadena proteica presenta un extremo carboxiterminal hidrofílico y se convierte en una molécula de exportación.
(Los linfocitos B, antes o después de ser activados, también pueden experimentar un cambio de clase, variando el tipo de cadena pesada de sus inmunoglobulinas. Por ejemplo, pueden cambiar de tipo M a tipo G. Estos cambios de clase no obedecen a un diferente procesamiento del ARNm, sino a la expresión de distintos segmentos de los genes de las cadenas pesadas. No obstante, nunca cambian su especificad antigénica. Ésta se define tempranamente y se conserva de por vida.)
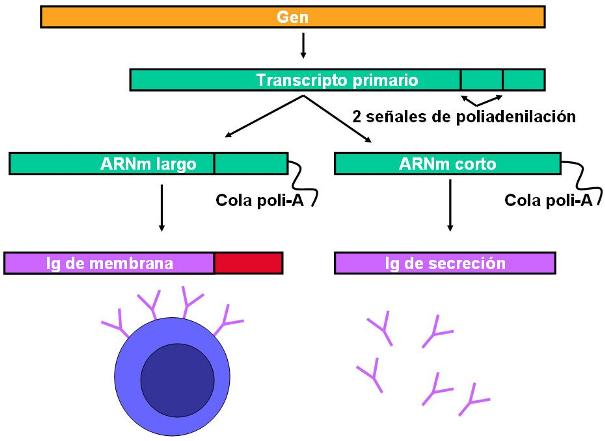 |
Las inmunoglobulinas circulantes participan en el ataque al patógeno específico de varias formas. Dado que la defensa es ejercida directamente por estas moléculas solubles, se habla de una inmunidad humoral (humor: líquido corporal).
| Complejo Ag-Ac |
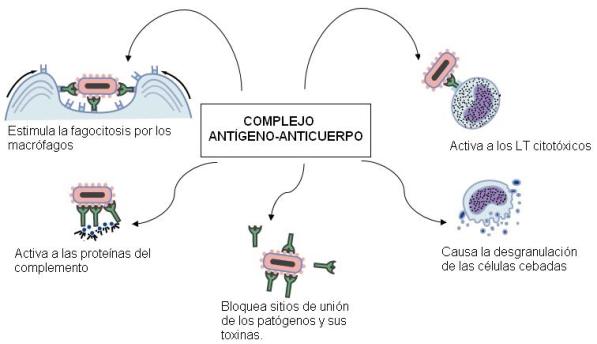
| Las inmunoglobulinas se unen a los antígenos en la superficie de los patógenos, formando los complejos Ag-Ac. Estos complejos combaten a los patógenos por los siguientes mecanismos: |
• Favorecen la fagocitosis por los macrófagos. Ésta es más eficiente cuando el patógeno está recubierto por antígenos (opsonización).
• Activan al sistema del complemento.
• Inactivan toxinas bacterianas.
• Bloquean moléculas de membrana que los patógenos intracelulares utilizan para ingresar a las células huésped.
• Activan a las células cebadas, que se desgranulan, iniciando la respuesta inflamatoria.
• Activan a las células citotóxicas. |
Respuesta inmune primaria y respuesta inmune secundaria Regresar
Cuando un patógeno ingresa al organismo por primera vez, éste se defiende mediante una respuesta inmunitaria primaria, ya sea celular o humoral, al cabo de un retraso de varios días. La segunda exposición al antígeno genera una respuesta secundaria, de mayor intensidad y con menor período de retraso que la primaria. Esto significa que el cuerpo ha “recordado” al antígeno y se verifica aunque entre una y otra respuesta hayan transcurrido muchos años.
La respuesta secundaria siempre es más efectiva que la primaria. Por esta razón es que pueden llegar a desarrollarse los síntomas de enfermedad la primera vez, pero no la segunda.
La memoria inmunológica se debe al aumento en el número de linfocitos después de su activación (expansión clonal) y a su diferenciación. Los linfocitos que se diferencian en células de memoria viven muchos años y ante una segunda exposición se activan con mayor celeridad. Además, al ser activados, pueden diferenciarse tanto en células efectoras como en células de memoria.
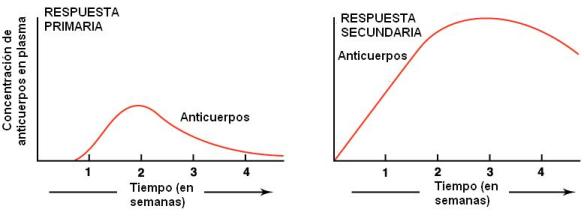 |
Inmunización artificial Regresar
La inmunización artificial es la que se consigue mediante las vacunas y los sueros. La inmunidad alcanzada a través de la vacunación es activa, en tanto la conferida por los sueros es inmunidad pasiva.
Una vacuna consiste en una preparación de antígenos que han sido modificados para que pierdan patogenicidad, pero conserven su poder de estimular la respuesta inmune. Esto se logra con gérmenes muertos o atenuados, o bien separando y seleccionando de los patógenos determinadas moléculas que funcionan como antígenos.
La vacuna produce en el individuo una respuesta inmune primaria, sin causar enfermedad. Después de un tiempo, el individuo ha generado células de memoria. Si tiene una exposición posterior al antígeno “real”, desarrolla la respuesta inmune secundaria, sin llegar a enfermar.
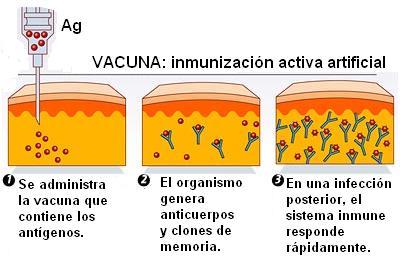 |
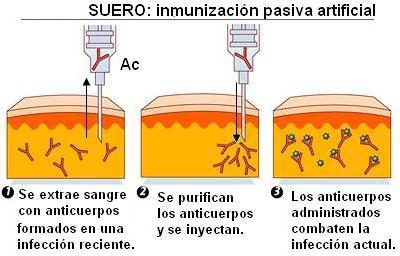
Los sueros se obtienen de animales que han sido inoculados con un determinado germen o su toxina. Estos animales reaccionan con una respuesta primaria. Los anticuerpos son extraídos del suero y administrados al individuo que debe ser protegido.
La inmunización mediante sueros es pasiva, pues quien la recibe no desarrolla ninguna respuesta inmune y no forma células de memoria. Además, la protección es más fugaz, ya que después de un breve lapso los anticuerpos son degradados. La ventaja de los sueros es que actúan inmediatamente, por lo que se los usa cuando la infección ya ocurrió o el riesgo de la misma es alto.
Las vacunas contienen antígenos |
Los sueros contienen anticuerpos |
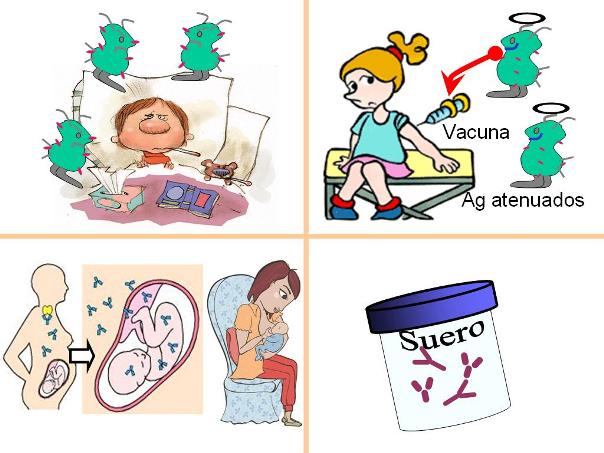
Inmunidad activa y pasiva Regresar
| Inmunidad adquirida |
Natural |
Artificial |
Activa: el organismo tiene una primera exposición al antígeno y produce una respuesta inmune primaria.Es duradera y genera memoria. |
Se adquiere por exposición a un patógeno ambiental. |
Se adquiere mediante la inoculación del antígeno (vacuna). |
Pasiva: el organismo recibe anticuerpos. No hay respuesta primaria. Es inmediata, fugaz y no genera memoria. |
La adquieren el feto o el recién nacido mediante el pasaje de anticuerpos maternos a través de la placenta o de la lactancia. |
Se adquiere mediante la administración de un suero rico en anticuerpos.
|
| |
Natural |
Artificial |
| Activa |
 |
| Pasiva |
|
|