|
Capítulo 10: Sistema Locomotor
Órganos del sistema locomotor Regresar
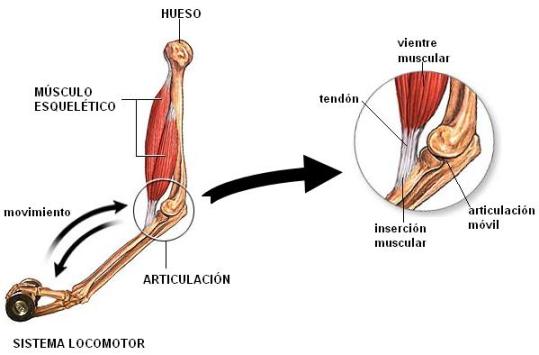
El sistema locomotor está formado por el esqueleto, un conjunto de huesos articulados, y los músculos esqueléticos, que se insertan en los huesos.
Los huesos proporcionan el sostén del cuerpo y brindan protección a otros órganos. También son el punto de apoyo para los músculos.
Las articulaciones son las estructuras que conectan a los huesos entre sí. La mayoría de ellas permite la movilidad de los huesos que ponen en contacto; otras son inmóviles.
Los músculos esqueléticos, bajo el control directo del sistema nervioso, son los responsables del movimiento.
Esqueleto Regresar
El esqueleto humano está formado por algo más de 100 huesos, algunos de ellos pares y otros impares.
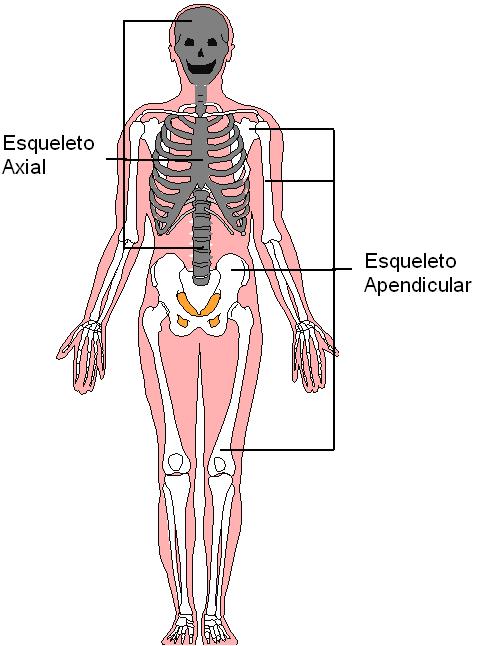
El esqueleto puede ser dividido en dos partes: axial y apendicular.
El esqueleto axial (de “axis”: eje) se encuentra en la línea media del cuerpo. Incluye el cráneo, la columna vertebral y la caja torácica.
El esqueleto apendicular comprende el esqueleto de las extremidades y las cinturas óseas, los huesos que conectan a las extremidades con la columna vertebral.
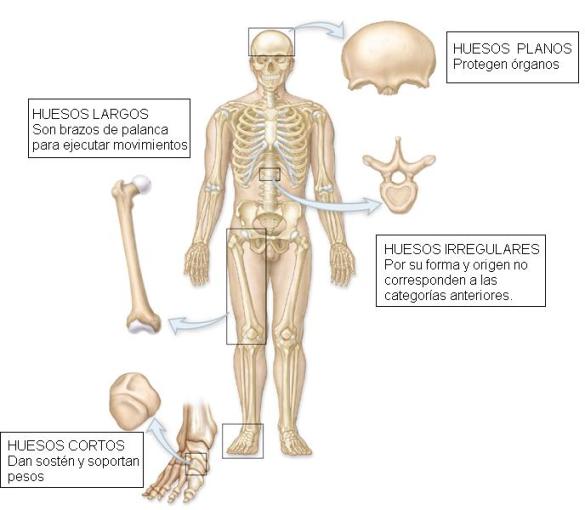
Clasificación de los huesos según sus dimensiones Regresar
Tejido óseo Regresar
Los huesos están formados primordialmente por tejido óseo, aunque éste es acompañado por tejido conectivo propiamente dicho y por tejido cartilaginoso.
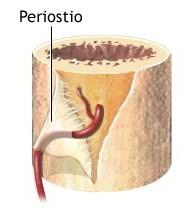 |
El tejido conectivo forma el periostio y el endostio, membranas que revisten las superficies externa e interna de los huesos, respectivamente.
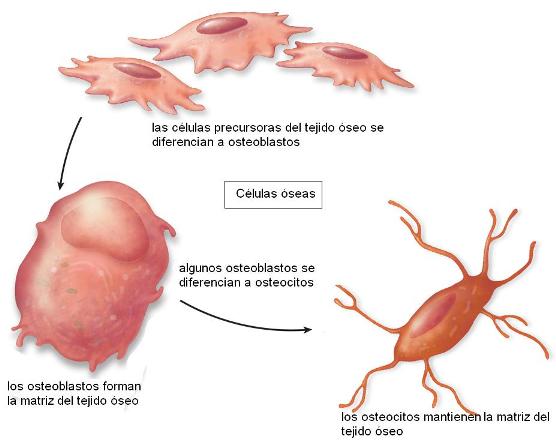
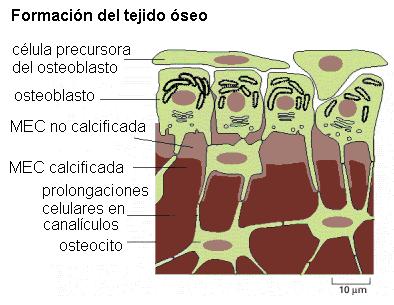
Las células formadoras del tejido óseo son los osteoblastos. Los osteoblastos segregan los componentes de la matriz extracelular (MEC); son células con un importante desarrollo del REG. La matriz extracelular recibe el nombre de sustancia osteoide y contiene proteoglucanos, glicoproteínas y abundantes fibras colágenas.
Los osteoblastos depositan sobre las fibras colágenas las sales de hidroaxipatita (fosfato de calcio) que mineralizan el hueso, otorgándole rigidez. Gran parte del calcio y del fosfato del organismo se encuentran en el tejido óseo.
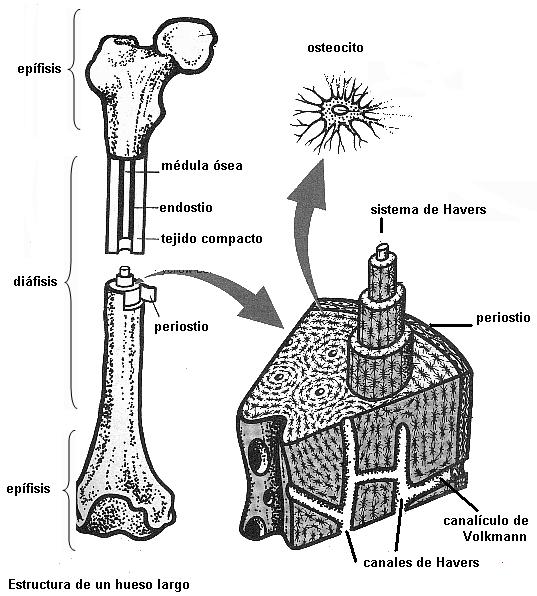
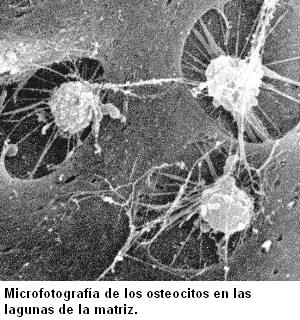
Cuando las células quedan completamente rodeadas por la MEC, reciben el nombre de osteocitos. Éstas presentan un cuerpo central, donde se ubican el núcleo y la mayor parte del citoplasma, y prolongaciones delgadas que les dan un aspecto araneiforme (forma de araña).
Las prolongaciones de los osteocitos se extienden por el interior de canalículos que quedan excavados en la sustancia intercelular y, a través de los mismos, distintos osteocitos pueden conectarse entre sí.
 |
 |
|
A medida que se produce el depósito de MEC alrededor de las células, durante la formación del hueso, quedan constituidas las laminillas de tejido óseo.
Las laminillas óseas pueden relacionarse unas con otras de diferente manera, determinando así dos variedades de tejido óseo: esponjoso y compacto.
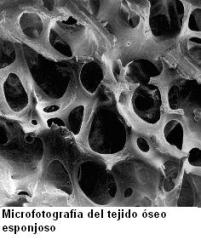 |
En el tejido óseo esponjoso, las laminillas se disponen formando trabéculas, que siguen distintas direcciones del espacio, en un entramado con huecos, semejante a una esponja. En los espacios del tejido esponjoso se aloja la médula ósea, un tejido blando cuya función, la hematopoyesis, consiste en la formación de células sanguíneas.
En el tejido óseo compacto, cada laminilla de tejido se superpone con sus vecinas sin dejar espacios, en un íntimo contacto.
La arquitectura de las laminillas depende de la disposición de las fibras colágenas en la MEC.
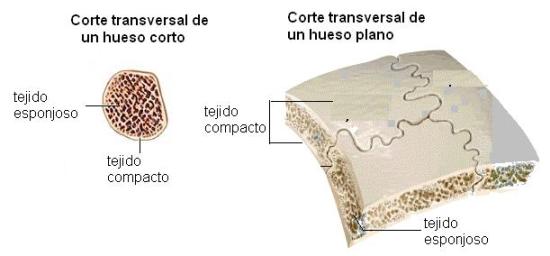
Las dos variedades de tejido óseo se encuentran formando parte de los huesos cortos, planos y largos, distribuidas según se detalla en el siguiente cuadro:
Tipo de Hueso |
Ubicación del Tejido Óseo Compacto |
Ubicación del Tejido Óseo Esponjoso |
| Corto |
Se dispone en la periferia del hueso, en laminillas paralelas a la superficie de cada una de las caras. |
Ocupa el interior del hueso. |
| Plano |
Forma dos tablas, una en cada cara del hueso. |
Ocupa el espacio central, comprendido entre las dos tablas de hueso compacto. Se lo denomina diploe. |
| Largo |
Epífisis(Extremo) |
La epífisis es similar a un hueso corto. El tejido compacto se dispone en la superficie de la epífisis. |
Ocupa el interior de la epífisis. |
| Diáfisis (cuerpo) |
Reviste la superficie externa de la diáfisis y delimita el canal medular, ubicado en el centro.Entre ambas capas se ubican los sistemas de Havers. (Ver aclaración). |
Entre los sistemas de Havers. |
 |
La diáfisis o cuerpo de los huesos largos presenta un revestimiento de tejido óseo compacto, formado por laminillas concéntricas dispuestas a modo de un manguito.
Internamente, otro manguito de tejido óseo compacto forma el límite del canal medular excavado en la parte central del hueso. El canal medular contiene médula ósea.
El espesor de la diáfisis, comprendido entre ambos manguitos de tejido compacto, está ocupado por los sistemas de Havers u osteones. Cada sistema de Havers es un cilindro formado por varias laminillas óseas concéntricas, que contiene un vaso sanguíneo en su parte central. Canalículos transversales conectan los sistemas de Havers entre sí a distintas alturas, permitiendo la comunicación entre los vasos sanguíneos que nutren al hueso.
Osificación Regresar
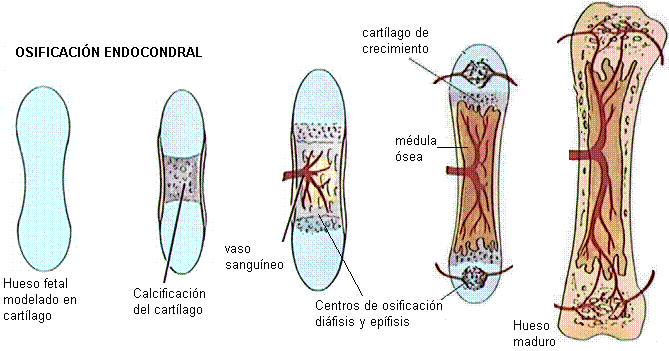
La osificación es el proceso de formación de los huesos, que comienza en la vida intrauterina. Hay dos tipos de osificación: intramembranosa y endocondral.
En la osificación intramembranosa, los huesos se forman cuando las células fetales precursoras de los tejidos conectivos (mesénquima) se transforman en tejido óseo. Dicha transformación se inicia en ciertos puntos del hueso llamados centros de osificación, y a partir de ellos el proceso se extiende al resto del hueso. La osificación intramembranosa ocurre, por ejemplo, en los huesos del cráneo.
Otros, como los huesos largos, se modelan primero en cartílago. Éste es posteriormente reabsorbido y reemplazado por tejido óseo. Esta forma de osificación se denomina endocondral.
Ya osificados, los huesos largos conservan un sector de cartílago en la zona de unión de la diáfisis con las epífisis: el cartílago de crecimiento. Éste no se reemplaza por tejido óseo sino hasta los 20 ó 25 años; entretanto, la división celular del cartílago de crecimiento permite el crecimiento en longitud del hueso.
 |
Crecimiento y remodelación óseos Regresar
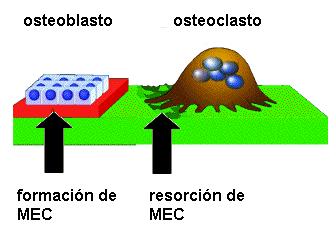
El tejido óseo es metabólicamente muy activo. Los huesos crecen durante la infancia y la adolescencia y realizan una continua actividad de remodelación a lo largo de la vida. La remodelación está influida por la actividad física desarrollada. El crecimiento y la remodelación de los huesos resultan de la acción combinada de los osteoclastos y los osteoblastos.
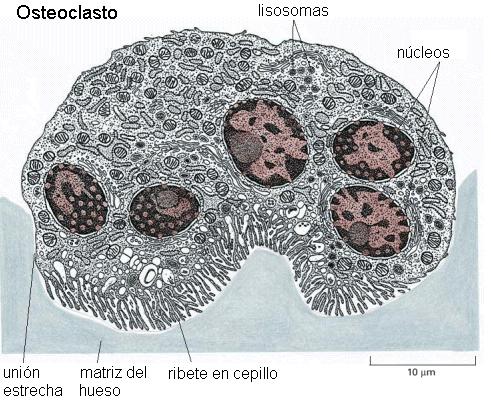
Los osteoclastos son células de gran tamaño, multinucleadas, formadas por la fusión de monocitos provenientes de la médula ósea y con un elevado contenido de enzimas lisosómicas.
Cuando un hueso va a ser remodelado, los osteoclastos aparecen sobre la superficie del mismo. Interactúan con la MEC a través de una superficie secretora plegada (ribete en cepillo) liberando ácidos y enzimas que provocan la degradación de sus componentes. Este proceso se denomina resorción ósea.
El crecimiento de los huesos requiere resorción de algunas partes a cargo de los osteoclastos, y formación de nueva matriz, a cargo de los osteoblastos.
 |
Después de la adolescencia, la remodelación del hueso continúa. En pequeñas placas dentro del hueso se van alternando procesos de resorción y formación de tejido. Sin embargo, ya no hay aumento de la masa ósea, porque ambos procesos se compensan. A partir de los 35-40 años, la resorción se hace predominante, causando una pérdida gradual de masa ósea.
Después de una fractura, el hueso se autorrepara repitiendo el proceso embrionario original. Para ello, requiere un modelo de cartílago en la zona de fractura. Las células cartilaginosas se diferencian a partir del periostio. El cartílago es reemplazado luego por el trabajo conjunto de osteoclastos y osteoblastos.
Articulaciones Regresar
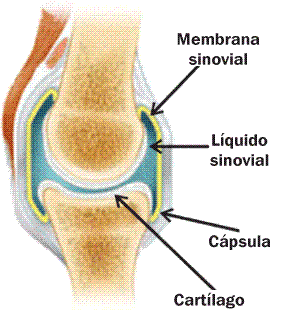
Las articulaciones son conjuntos de partes duras y blandas que conectan a dos o más huesos.
Las partes duras de la articulación corresponden a las superficies articulares de los huesos.
Las partes blandas incluyen estructuras conectivas, como las cápsulas y los ligamentos, formados por tejido conectivo denso. Éstos sostienen a la articulación, ayudando a mantener en su lugar las superficies óseas.
También forman parte de las articulaciones distintos tipos de cartílagos. Por ejemplo: el cartílago de revestimiento, que protege a las superficies óseas en algunas articulaciones, o los cartílagos articulares, como los meniscos de la articulación de la rodilla.
Las membranas sinoviales contienen un líquido seroso que favorece el deslizamiento de los huesos.
Si bien la mayoría de las articulaciones están dotadas de movilidad, existen articulaciones inmóviles. Estas últimas reciben el nombre de suturas y se encuentran entre los huesos de la cabeza.
Las articulaciones más móviles se hallan en las extremidades.
 |
Tejido cartilaginoso Regresar
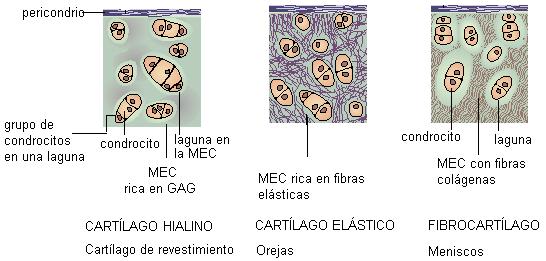
El tejido cartilaginoso está compuesto por células llamadas condroblastos. Éstas segregan la matriz extracelular, que contiene proteoglucanos, fibras elásticas y fibras colágenas, elementos cuyas proporciones varían dando origen a los diferentes tipos de cartílago: hialino, elástico y fibroso.
A medida que los condroblastos quedan rodeados por la MEC, se separan unos de otros y permanecen encerrados en lagunas (condroplastos); al llegar a este estado de madurez los condroblastos se convierten en células menos activas llamadas condrocitos. Cuando las células se dividen, las células hijas permanecen dentro de la misma laguna formando un grupo.
El tejido cartilaginoso está rodeado por una membrana conectiva denominada pericondrio, en la cual pueden diferenciarse nuevos condroblastos que permiten el crecimiento del cartílago.
El tejido cartilaginoso no posee irrigación propia. Los nutrientes deben difundir hasta las células cartilaginosas desde los vasos sanguíneos cercanos, atravesando la sustancia intercelular.
El cartílago es un tejido de sostén, cuya consistencia es más blanda que la del hueso, pues no está mineralizado como aquél, pero más rígida que la del tejido conectivo propiamente dicho. Sus características lo hacen apto para cumplir funciones protectoras, amortiguadoras y para brindar un sostén flexible.
En el siguiente esquema se muestran las variedades del tejido cartilaginoso y sus respectivas ubicaciones.
 |
Músculos esqueléticos Regresar
Los músculos esqueléticos, situados entre el tejido adiposo subcutáneo y los huesos, se fijan a estos últimos, permitiéndonos mantener la postura corporal y ejecutar infinidad de movimientos.
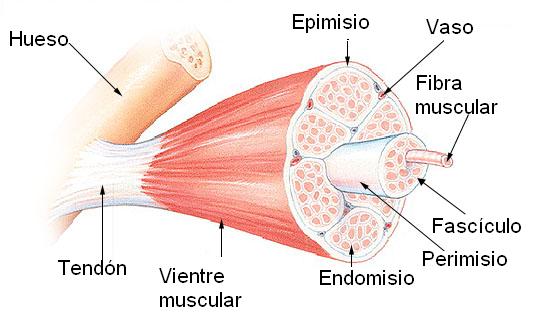
Los músculos poseen una porción central, el vientre, donde se localiza el tejido muscular estriado, principal tejido de estos órganos y responsable de sus propiedades funcionales.
Las células del tejido muscular, llamadas fibras, por su forma cilíndrica alargada, aparecen acompañadas por tejido conectivo. Éste forma vainas que envuelven a cada fibra muscular, a manojos de fibras musculares y al vientre en su totalidad. Dichas envolturas reciben, respectivamente, los nombres de endomisio, perimisio y epimisio.
La vaina conectiva se prolonga desde los extremos del vientre constituyendo los tendones, cordones fibrosos resistentes, cuya función es insertar el músculo en los huesos.
Fibra muscular estriada Regresar
Las células del tejido muscular estriado esquelético son largas y delgadas y reciben el nombre de fibras. Una fibra estriada proviene de la fusión de varias células, ocurrida durante la vida embrionaria. Los núcleos de las células precursoras se conservan, ubicándose en la periferia del citoplasma, por debajo de la membrana plasmática.
Las fibras estriadas son células con un alto grado de especialización morfológica, adaptadas para producir la contracción muscular. La especialización abarca principalmente al citoesqueleto, pero también a la membrana plasmática (llamada sarcolema, de sarco: carne y lema: membrana), al REL (llamado retículo sarcoplásmico) y a las mitocondrias (llamadas sarcosomas), que adquieren grandes dimensiones. Las mitocondrias proveen la energía para la contracción, en forma de ATP; por lo tanto, su número se incrementa de acuerdo con la actividad muscular.
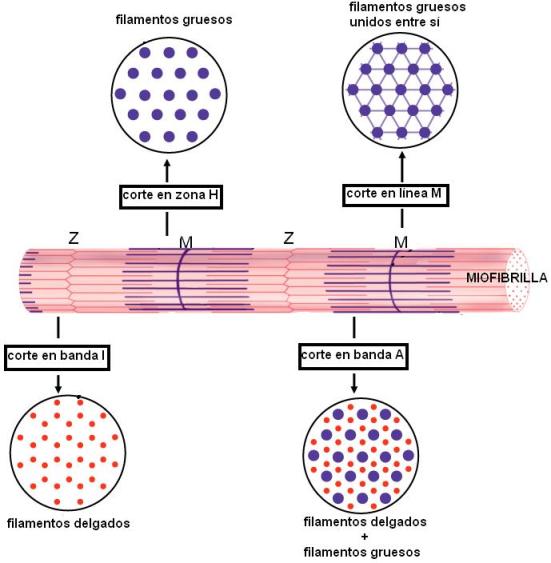
En el citoplasma de las fibras musculares se encuentran las miofibrillas. Cada miofibrilla es una estructura cilíndrica que se extiende a lo largo de toda la célula muscular. Las miofibrillas están formadas por la repetición de las unidades contráctiles: los sarcómeros. Dicha repetición es lo que le confiere a la fibra muscular su aspecto estriado (estría: raya).
Cada sarcómero consta de dos tipos de filamentos: los filamentos gruesos, constituidos por miosina, y los filamentos delgados, compuestos por actina y otras proteínas. Los filamentos gruesos y delgados se superponen parcialmente y el deslizamiento de unos sobre otros es lo que causa la contracción o acortamiento del sarcómero.
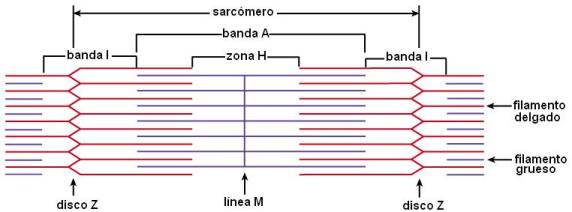 |
En un sarcómero se distinguen la banda I y la banda A (I proviene de “isótropa” y A de “anisótropa”. Estos términos se relacionan con el tipo de imagen que ofrecen las bandas al microscopio de luz polarizada.)
La banda I está formada solamente por filamentos delgados. Éstos están unidos al disco Z.
La banda A presenta superposición de filamentos delgados y gruesos en su mayor parte, excepto en su sector central, donde solamente hay filamentos gruesos; ese sector se denomina zona H. La línea media de la zona H, línea M, corresponde a la unión entre los filamentos gruesos.
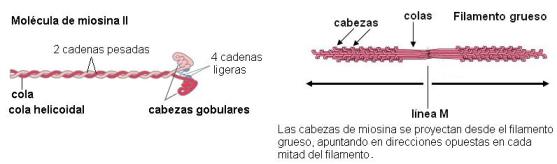
Los filamentos gruesos están formados por miosina II. Cada molécula de miosina II es una proteína de 6 cadenas polipeptídicas, que se unen en una estructura con dos cabezas globulares y una cola helicoidal. Las colas de distintas miosinas se empaquetan entre sí, formando un filamento bipolar, del cual sobresalen las cabezas. Las cabezas de la mitad del filamento tienen dirección opuesta a las de la otra mitad.
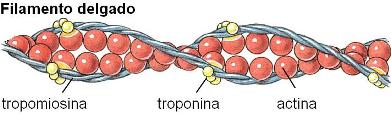
En los filamentos delgados hay microfilamentos de actina, y otras dos proteínas: tropomiosina y troponina. La tropomiosina consta de dos hélices que se ubican en el surco del microfilamento de actina. La troponina es un complejo de tres pequeñas proteínas.
Igual que los filamentos gruesos, los filamentos delgados están polarizados. Los extremos más de los filamentos delgados se fijan al disco Z y los extremos menos se orientan hacia la línea media del sarcómero.
Muchas más proteínas forman parte del sarcómero; algunas de ellas son estructurales y otras, reguladoras. Estas proteínas mantienen la organización del sarcómero. Retículo sarcoplásmico Regresar
El retículo sarcoplásmico está formado por una red de túbulos anchos, longitudinales, que envuelven a las miofibrillas. A la altura de la línea Z o en la unión entre bandas A y bandas I, según el tipo de célula, los túbulos se amplían dando lugar a las cisternas terminales. Entre dos cisternas terminales consecutivas se extiende el túbulo T, que es una invaginación de la membrana plasmática. Dos cisternas terminales y el túbulo T interpuesto conforman una tríada.
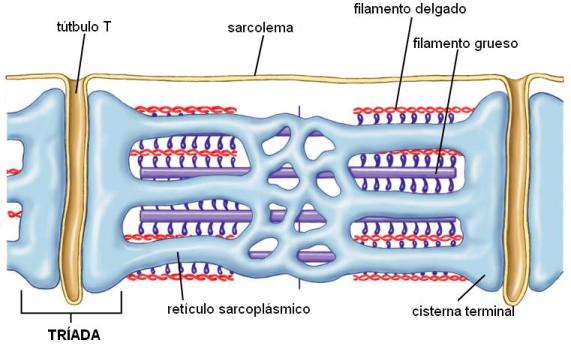 |
El retículo sarcoplásmico es un reservorio de calcio iónico (Ca2+). Dentro del retículo, la concentración de este ión es muy alta, comparada con la concentración citoplasmática.
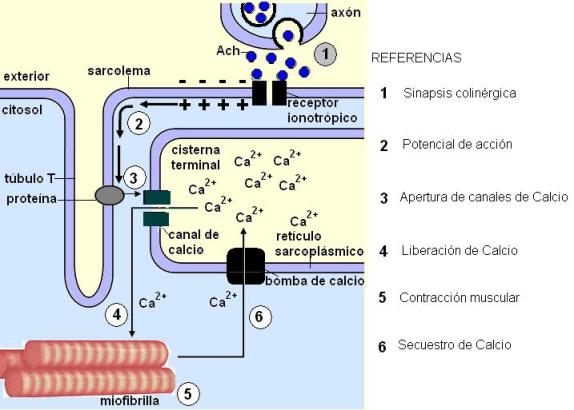
La neurona motora y la célula muscular establecen contacto en la llamada unión neuromuscular o placa motora, donde se realiza una sinapsis química mediada por acetilcolina. Los receptores para acetilcolina (receptores colinérgicos) de la célula muscular son canales de Na+ regulados por ligando. Cuando se produce la sinapsis entre la neurona motora y la membrana de la célula muscular, el Na+ ingresa a través de los canales y la membrana de la célula muscular se despolariza. El potencial de acción se transmite a la profundidad de la célula a través de los túbulos T.
Ante la llegada del potencial de acción, proteínas membranares de los túbulos T cambian su conformación y abren los canales de calcio de las cisternas terminales.
El repentino aumento en la concentración de Ca2+ citosólico dispara la contracción del sarcómero.
De esta forma, el retículo sarcoplásmico actúa como un enlace entre la llegada de la orden motora, transmitida en la unión neuromuscular, y la respuesta de la contracción muscular.
El Ca2+ es rápidamente removido del citosol por la acción de una bomba específica ubicada en las membranas del retículo. El descenso en la concentración de Ca2+ citosólico conduce a la relajación muscular.
Mecanismo de la contracción muscular Regresar
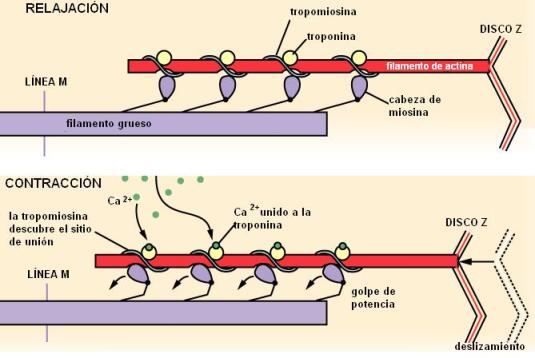
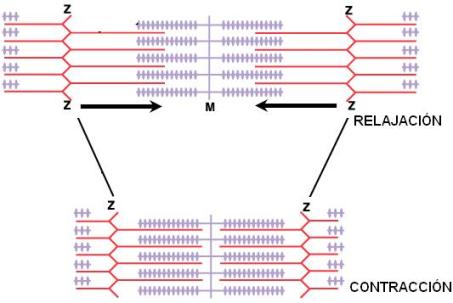
La contracción muscular se debe a un acortamiento del sarcómero producido por el deslizamiento de los filamentos delgados sobre los filamentos gruesos. Este deslizamiento tiene dirección opuesta en cada mitad del sarcómero, de manera que los discos Z de un mismo sarcómero se acercan entre sí. Durante la contracción, prácticamente se borra la banda H, a causa de la mayor superposición de filamentos delgados y gruesos.
 |
La contracción muscular es un fenómeno cíclico que implica la formación y ruptura de puentes cruzados entre las cabezas de miosina que se proyectan desde los filamentos gruesos y las moléculas de actina que forman los filamentos delgados.
Por cada puente formado resulta hidrolizada una molécula de ATP y la longitud del desplazamiento causado corresponde a la de una unidad de actina globular (5,3 nm). La velocidad de la contracción es de 15 micrómetros/seg.
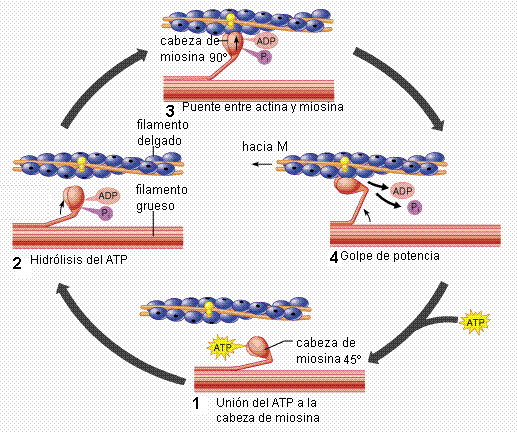
En el estado de rigor o rigidez, la actina está unida a la cabeza de miosina en un ángulo de 45º. Cuando se inicia la contracción (1), la miosina queda libre para unirse a una molécula de ATP, separándose de la actina.
La cabeza de miosina posee actividad ATPasa y causa la hidrólisis del ATP (2); sin embargo, los productos de la hidrólisis, ADP y P, permanecen unidos a la miosina.
La miosina forma un puente con otra molécula de actina, situada hacia el extremo más del filamento delgado (3). El ángulo de este puente es de 90º. Una vez unida a la actina, la miosina libera el P. La liberación del P provoca un cambio conformacional en la miosina, produciendo ésta un “golpe de potencia” (4).
El golpe de potencia es el causante del deslizamiento del filamento delgado, en dirección hacia la línea M. Esto es lo que ocasiona el acortamiento del sarcómero. Después del golpe de potencia, la miosina libera el ADP y el ciclo recomienza.
 |
Control de la contracción muscular mediado por el calcio Regresar
El control de la contracción mediado por el calcio involucra a las proteínas reguladoras del filamento delgado: la tropomiosina y la troponina.
En el estado de relajación, la tropomiosina bloquea el sitio de unión para la miosina sobre la actina G. Si en ese momento se produce un aumento de la concentración de Ca2+ citosólico (como ocurre cuando llega un estímulo por la neurona motora), éste es fijado por la troponina. La troponina unida al calcio desplaza a la tropomiosina dentro del filamento delgado. La nueva posición de la tropomiosina deja libre el sitio de unión de la actina para la miosina. Entonces éstas establecen una unión fuerte y sobreviene el golpe de potencia.
|
|