|
Capítulo 9: Sistema nervioso
Organización general del sistema nervioso Regresar
El sistema nervioso es el principal sistema de relación y control que han desarrollado los animales y que ha alcanzado su máxima expresión en los mamíferos, especialmente en el hombre.
Es el encargado de captar la información proveniente del medio externo y del medio interno, centralizar y procesar dicha información y elaborar respuestas coordinadas que no solo permitan armonizar las funciones internas sino también ajustar el organismo al ambiente que lo rodea.
Las respuestas y cambios producidos por el sistema nervioso son rápidos, ocurren en fracciones de segundo.
El sistema nervioso es muy plástico, pues además de coordinar funciones innatas, también almacena información (memoria) y modela sus funciones a partir de la experiencia, posibilitando las modificaciones del comportamiento a las que llamamos aprendizaje.
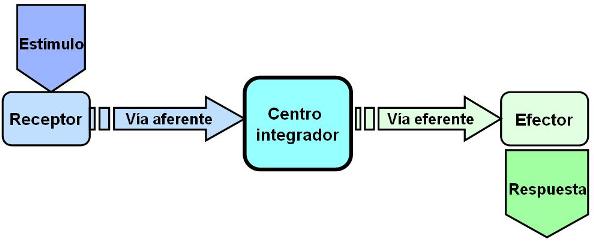
En el funcionamiento del sistema nervioso se interrelacionan los siguientes elementos:
• Estímulo. Es cualquier cambio físico o químico producido en el medio externo o en el medio interno, que el sistema nervioso pueda detectar. Por ejemplo: luz, temperatura, presión, sonido, osmolaridad.
• Receptor. Es la estructura especializada para captar un determinado tipo de estímulo. Por ejemplo: fotorreceptores de la retina, receptores de dolor en la piel, osmorreceptores en los vasos sanguíneos.
• Vía sensitiva o aferente. Es la estructura por la cual la información entrante o aferente, también llamada sensitiva, viaja desde el receptor hasta un centro nervioso.
• Centro integrador. Es el órgano del sistema nervioso donde se centraliza información aferente y se elabora la respuesta adecuada.
• Vía motora o eferente. Es la estructura por la cual viaja la información necesaria para producir una respuesta, desde el centro integrador hasta el órgano efector.
• Órgano efector. Recibe la información eferente y efectúa una acción en consecuencia. Los órganos efectores son músculos o glándulas.
• Respuesta. Es la acción ejecutada por el órgano efector.
 |
Divisiones anatómica y funcional del sistema nervioso Regresar
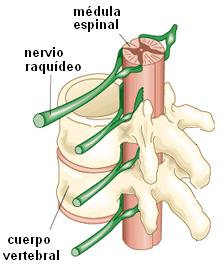
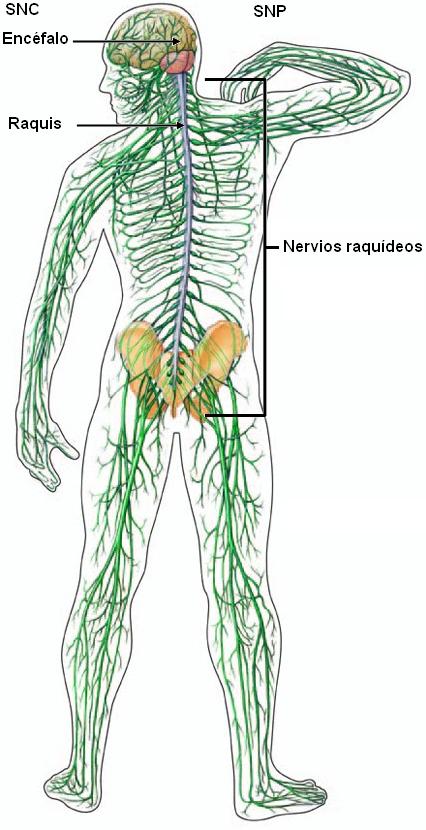
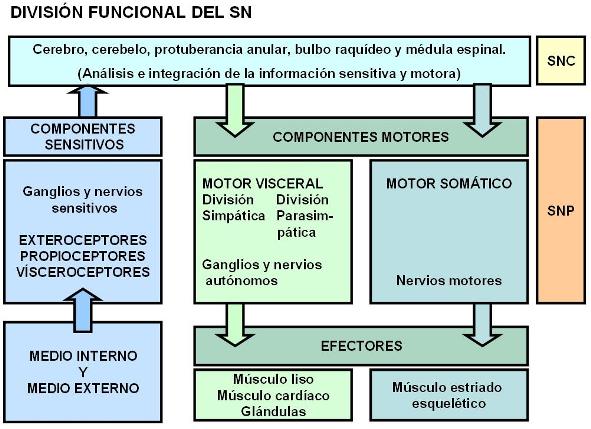
Desde el punto de vista anatómico, el sistema nervioso (SN) se divide en sistema nervioso central (SNC) y sistema nervioso periférico (SNP). El SNC está formado por los órganos que, en conjunto, forman el encéfalo y se encuentran protegidos por la caja craneana: cerebro, cerebelo, protuberancia anular y bulbo raquídeo; más la médula espinal o raquis , protegida por la columna vertebral. Todos ellos están envueltos por tres membranas: duramadre, aracnoides y piamadre, colectivamente llamadas meninges.
Órganos del SNC |
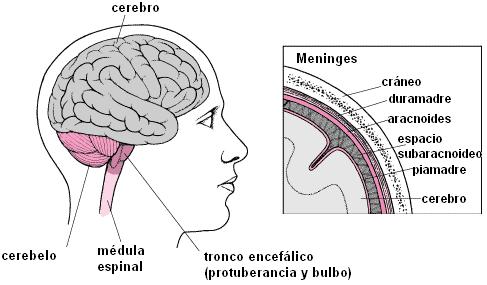 |
|
El SNP está formado por los nervios, que conectan el SNC con los órganos, y los ganglios nerviosos. Los nervios que nacen del encéfalo se denominan nervios craneales y son 12 pares. Los nervios que nacen de la médula espinal son los nervios raquídeos, en total 31 pares.
|
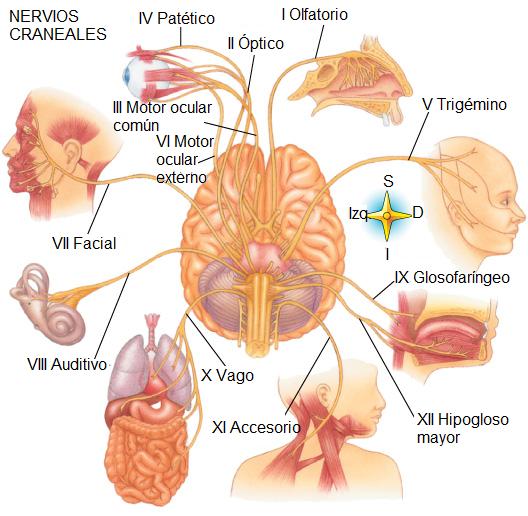 |
Desde el punto de vista funcional, el SN ha sido dividido en un sistema nervioso de la vida de relación o somático, que nos conecta con el entorno, y un sistema nervioso autónomo, que coordina las funciones viscerales. Sin embargo, esta división no tiene un correlato anatómico exacto, ya que un mismo nervio puede conducir al mismo tiempo información procedente del exterior o del interior del cuerpo o inervar tanto estructuras somáticas como viscerales. Además, todas las aferencias o información sensitiva se interconectan a nivel del SNC.
Los estímulos externos e internos son captados en distintos tipos de receptores especializados: exteroceptores, propioceptores y vísceroceptores.
La información procedente del medio externo es captada por los exteroceptores ubicados en la piel, que registran presión, tacto, dolor, frío y calor y por los órganos de los sentidos especiales, como las papilas gustativas de la lengua, los receptores olfatorios de la mucosa nasal, los receptores del oído interno y los fotorreceptores (conos y bastones) de la retina.
La información procedente de los músculos y articulaciones es captada por los propioceptores.
La información procedente de las vísceras es recibida en los vísceroceptores.
Toda esta información llega por medio de vías aferentes, que transcurren en parte dentro de los nervios y en parte dentro del SNC, a algún centro sensitivo ubicado en el SNC.
En el SNC se integra toda la información recibida y se establece la conexión con los centros motores, ubicados también dentro del SNC. Desde los centros motores parte información eferente o motora hacia los efectores. La información motora viaja en parte dentro del SNC y en parte por el interior de los nervios, hasta llegar a los efectores.
Los efectores son los músculos esqueléticos, el músculo liso visceral, el músculo estriado cardíaco y las glándulas.
Las vías eferentes o motoras que llegan a los músculos esqueléticos forman el sistema nervioso motor somático.
Las vías eferentes o motoras que llegan al músculo liso visceral, al músculo cardíaco y a las glándulas, forman el sistema nervioso autónomo o neurovegetativo, que como veremos más adelante, tiene dos divisiones: el sistema simpático y el parasimpático.
 |
Propiedades de las neuronas Regresar
El sistema nervioso está formado por el tejido nervioso. Éste consta de dos tipos celulares básicos: las neuronas y las células de la glía. El SN humano posee alrededor de un billón de neuronas y de 10 a 50 veces más células gliales.
Las neuronas son las células principales del tejido nervioso y las unidades anatómicas y funcionales del sistema. En las neuronas, la irritabilidad o capacidad de responder a estímulos, que es una propiedad universal en los seres vivos, alcanza su máximo desarrollo. La “información” que circula por el sistema nervioso lo hace a través de las neuronas. La información, también llamada “impulso nervioso” es de naturaleza eléctrica.
Las propiedades de las neuronas que les posibilitan cumplir su función son:
- Excitabilidad: generan un impulso nervioso ante un estímulo.
- Conducción: propagan el impulso nervioso a lo largo de su membrana.
- Transmisión: se comunican con otras neuronas o células efectoras, en las cuales desencadenan un nuevo impulso. La comunicación de una neurona con otra o con un efector se denomina sinapsis. |
Muchas veces el impulso nervioso se ha comparado con una corriente eléctrica. Así como la corriente eléctrica que viaja por los cables de un sistema eléctrico tiene siempre la misma naturaleza, el impulso nervioso que provoca un movimiento muscular es de la misma naturaleza que el que nos permite oír. La diferencia no está en el impulso, está en las conexiones.
El impulso nervioso no es exactamente una corriente eléctrica; ya aclararemos de qué se trata. Las conexiones del sistema nervioso son también más complejas que las de un sistema de cables eléctricos; sin embargo, la analogía es válida como una primera aproximación.
Estructura de la neurona Regresar
Las neuronas tienen diferentes formas y tamaños. No obstante, en todas las neuronas se pueden distinguir las mismas zonas, adaptadas a funciones específicas.
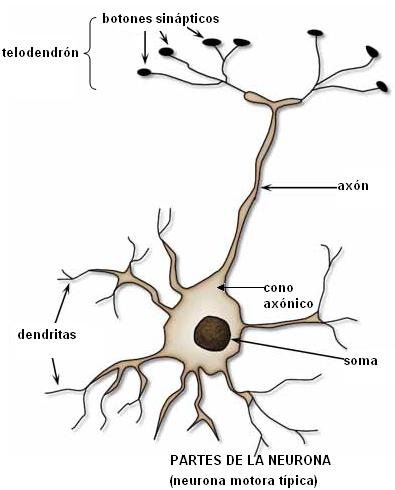
Una neurona (como se observa en el esquema de la neurona motora espinal) presenta un cuerpo neuronal y procesos o prolongaciones de dos tipos: dendritas y axón.
Las dendritas se extienden desde el cuerpo neuronal y se ramifican extensamente. Son, generalmente (aunque no siempre), la zona por donde una neurona recibe la información. En la superficie de las dendritas hay unas excrecencias llamadas espinas, donde se realizan los contactos sinápticos. El número y el tamaño de las espinas cambian, tanto durante el desarrollo embrionario como a lo largo de la vida; estos cambios están relacionados con la actividad de las neuronas y son la “huella” morfológica de cambios funcionales.
| |
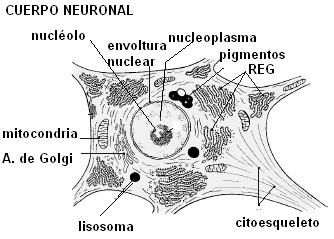 |
El cuerpo neuronal, también llamado soma o pericarion, contiene el núcleo. En el cuerpo neuronal se encuentran importantes cantidades de un material, la sustancia de Nissl, que corresponde a los ribosomas y al REG. También el aparato de Golgi se halla bien desarrollado. El pericarion concentra casi toda la actividad biosintética de la neurona y de esta actividad depende asimismo la mantención de las prolongaciones, cuya extensión supera ampliamente la del cuerpo celular.
El cono axónico es la región del soma de donde se origina el axón. El axón es una prolongación única, cuyo extremo o telodendrón se divide en ramas terminales, los botones sinápticos. En los botones sinápticos se acumulan las vesículas sinápticas, que almacenan los neurotransmisores. Éstos son señales químicas que participan en la comunicación intercelular o sinapsis. Cuando una neurona es excitada, el impulso nervioso se propaga hasta el axón y desde allí se liberan los neurotransmisores.
Tanto en la construcción como en el mantenimiento de su estructura, el citoesqueleto de las neuronas cumple un papel fundamental. Cuando las neuronas se originan, durante el desarrollo embrionario, migran hacia sus localizaciones definitivas valiéndose de los filamentos de actina. Una vez allí, comienzan a crecer sus axones y dendritas. Cada una de estas prolongaciones se extiende a partir de un cono de crecimiento que avanza gracias a las estructuras que forma la actina en su interior. El resto de la dendrita o el axón va detrás del cono de crecimiento. Los conos de crecimiento son guiados por señales químicas de la matriz extracelular o moléculas secretadas por otras células, que los atraen o repelen, determinando así la dirección del crecimiento. Los microtúbulos se organizan en el cono de crecimiento, reforzando la decisión direccional tomada por las estructuras ricas en actina en el frente de avance.
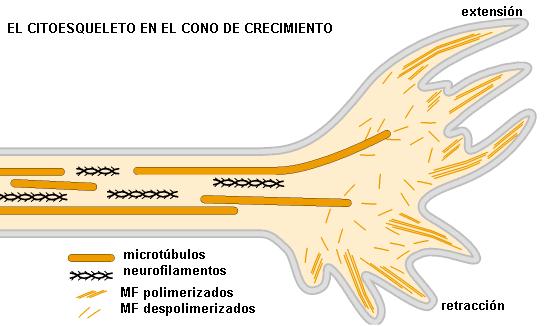 |
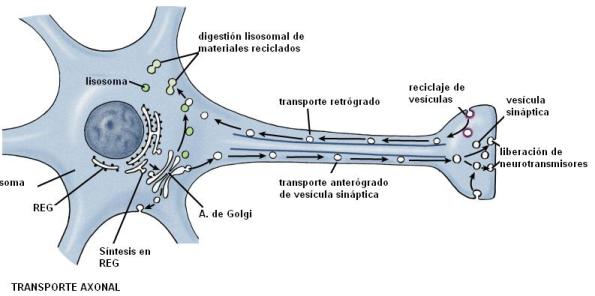
En los axones, los microtúbulos se disponen todos en la misma dirección (con sus extremos más hacia el telodendrón) formando haces que se van superponiendo y determinan una verdadera pista de transporte a lo largo del axón.
Sobre los microtúbulos, las proteínas motoras transportan vesículas sinápticas, mitocondrias y otras proteínas empacadas en vesículas, desde el cuerpo a la terminal axónica. Este tipo de transporte se denomina anterógrado. La quinesina es la proteína motora que se asocia a los microtúbulos en el transporte anterógrado.
También hay un transporte retrógrado, desde las terminales al soma, en el cual interviene la proteína motora dineína. De esta forma retornan al cuerpo celular algunas vesículas sinápticas para su reciclaje y ciertos materiales endocitados en el extremo del axón.
En las dendritas, los microtúbulos se ubican en paralelo, pero con sus polaridades mezcladas.
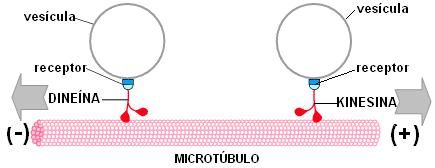 |
Las proteínas motoras asociadas a MT, quinesina y dineína, ejecutan movimientos como "pasos" sobre los MT, transportando vesículas y otras organelas, con gasto de energía. Los receptores son moléculas que median la unión de la organela a la proteína motora. La quinesina se desplaza hacia el extremo más y la dineína hacia el extremo menos del MT. |
Los otros componentes del citoesqueleto también contribuyen a la arquitectura y la función de las neuronas. Los filamentos de actina y sus proteínas motoras se encuentran por debajo de la membrana plasmática. Los neurofilamentos (filamentos intermedios propios del tejido nervioso) son el soporte estructural más importante de los axones.
Las neuronas son células metabólicamente muy activas. En el adulto, en condiciones normales, utilizan exclusivamente la glucosa como combustible y son muy sensibles a la hipoglucemia (descenso de la glucosa sanguínea), pues carecen de depósitos y dependen del suministro de glucosa a través de la sangre. Pese a representar tan solo el 2,5 % del peso corporal, consumen el 60% del total de glucosa y el 20% del total del oxígeno utilizados en condiciones de reposo. De allí que un bloqueo vascular, aun de pocos minutos, puede causar un daño irreversible en el cerebro.
Después del período embrionario, las neuronas ya no se dividen, permaneciendo en período Go de la interfase. La división celular haría que se perdiesen la información almacenada y los nuevos contactos funcionales que las neuronas desarrollan a lo largo de la vida.
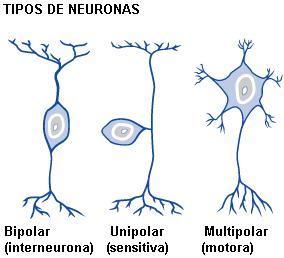
Tipos de neuronas Regresar
Desde el punto de vista morfológico, las neuronas se clasifican en:
• Unipolares: el axón y la única dendrita nacen del mismo polo celular.
• Bipolares: el axón y la dendrita nacen de polos opuestos del cuerpo celular.
• Multipolares: poseen un cuerpo estrellado, con numerosas dendritas.
Desde el punto de vista funcional, las neuronas son:
• Sensitivas,
• Motoras, o
• De asociación: también llamadas interneuronas, conectan a las neuronas sensitivas con las motoras.
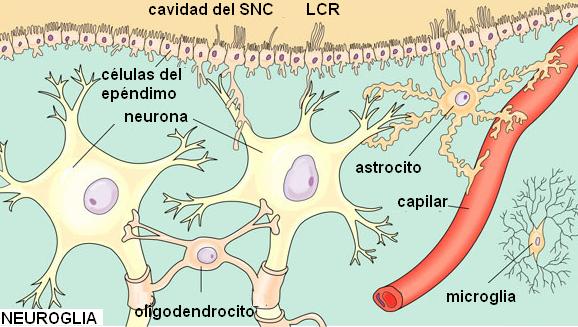
Neuroglia o células de la glía Regresar
Neuroglia significa “pegamento de la neurona”. Los cuerpos celulares, los axones y las dendritas de las neuronas están completamente rodeados por células gliales, las cuales son, como ya se mencionó, mucho más numerosas que las neuronas.
Existen cinco tipos de células gliales:
• Microglia: son pequeñas, con abundantes prolongaciones y capacidad fagocítica. La fagocitosis es un tipo de endocitosis consistente en la incorporación de partículas sólidas de gran tamaño. La microglia está emparentada con los macrófagos presentes en otros tejidos y procede de los monocitos. Las células de la microglia actúan como células de defensa y eliminando residuos.
• Astrocitos: sirven como soporte físico para las neuronas. Pueden proliferar formando tejido cicatricial cuando hay una lesión. Presentan prolongaciones con extremos dilatados (pies terminales) que rodean a los vasos sanguíneos. Los astrocitos inducen la formación de uniones estrechas en las células del endotelio capilar. En el SNC, el endotelio capilar es muy poco permeable. A diferencia del endotelio de otros tejidos, que es discontinuo o presenta poros, el endotelio dentro del SN actúa como una barrera. Las características del endotelio más la protección que ejercen los astrocitos son los responsables de lo que se ha dado en llamar la barrera hematoencefálica (BHE). La BHE aísla en gran medida al tejido nervioso de la circulación. Es muy eficaz para impedir el paso de sustancias hidrosolubles desde la sangre al tejido nervioso. También amortigua cambios bruscos que se producen en la concentración iónica del plasma e impide que neurotransmisores del SNC ingresen a la circulación fuera de él. Cuando la BHE se altera, por ejemplo en la acidosis (descenso del pH sanguíneo), isquemia (falta de oxígeno), hemorragias o infecciones, se produce la muerte neuronal.
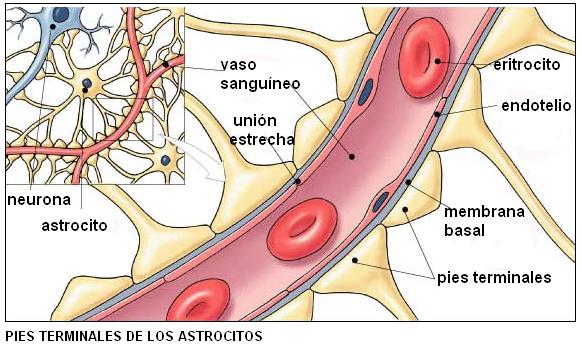 |
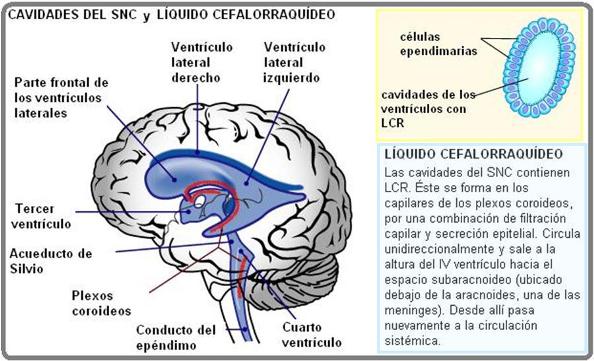
• Células ependimarias: en el interior de los órganos del SNC existen una serie de cavidades interconectadas, dentro de las cuales circula el líquido cefalorraquídeo (LCR). Las células ependimarias forman una monocapa que tapiza el interior de dichas cavidades.
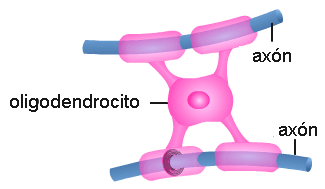
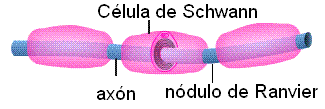
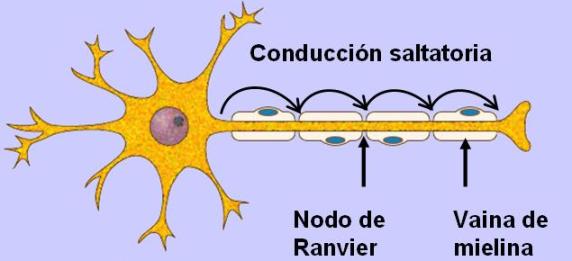
• Oligodendrocitos: son células ubicadas en el SNC. Un oligodendrocito posee varias prolongaciones laminares; cada prolongación envuelve el axón de una neurona. El axón envuelto toma el nombre de fibra nerviosa. Las membranas de los oligodendrocitos contienen mielina (un esfingolípido) que actúa como aislante, aumentando la velocidad de conducción del impulso nervioso. • Células de Schwann: se ubican en el SN periférico. Cada célula de Schwann envuelve al axón de una única neurona y forma a su alrededor una vaina celular. Entre el axón y la vaina celular se deposita una gruesa capa de mielina, la vaina de mielina. A lo largo de un axón hay varias células de Schwann; entre una célula y otra quedan zonas desprovistas de mielina. Las zonas del axón donde se interrumpe la vaina de mielina se denominan nódulos de Ranvier.
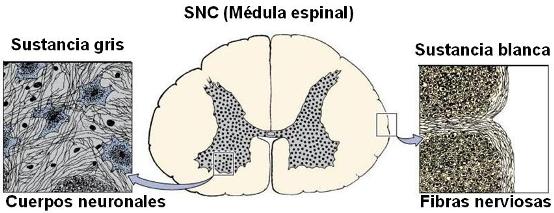
Sustancia gris y sustancia blanca Regresar
Dentro del SNC se distinguen la sustancia gris y la sustancia blanca. La sustancia gris está formada por los cuerpos neuronales y las prolongaciones neuronales que carecen de mielina (fibras amielínicas). La sustancia gris forma los núcleos del SNC y la corteza cerebral y cerebelosa.
La sustancia blanca está constituida por los axones con cubierta de mielina (fibras mielínicas) y forma las vías o tractos dentro del SNC.
Nervios y ganglios nerviosos Regresar
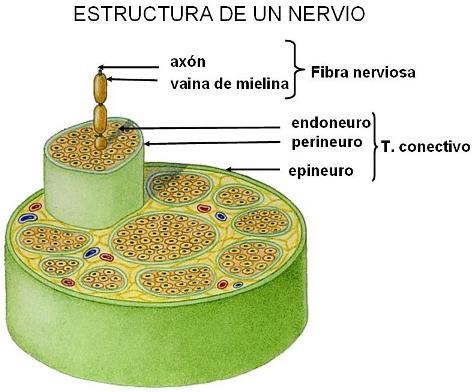
Los nervios son cordones formados por haces de fibras nerviosas que emergen del SNC. El nervio está envuelto por una vaina de tejido conectivo, que le proporciona sostén e irrigación.
En el trayecto de algunos nervios se acumulan cuerpos neuronales. Dichas estructuras se denominan ganglios nerviosos.
Ubicación de las neuronas en el SN |
Parte de la neurona |
SNC |
SNP |
Soma |
Sustancia gris (núcleos grises y corteza) |
Ganglio |
Prolongaciones |
Sustancia blanca (vías o tractos) |
Nervio |
Impulso nervioso Regresar
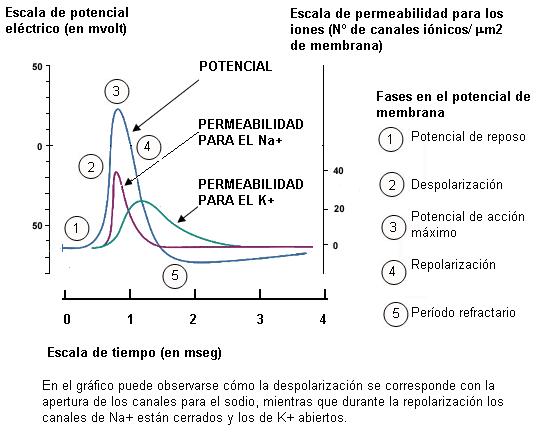
Durante el reposo, las neuronas se encuentran polarizadas. El continuo accionar de la bomba de Na+ y K+ (que extrae tres cationes por cada dos que introduce), sumado a la presencia de aniones no difusibles en el interior celular, determina que a través de las membranas se produzca un ligero desequilibrio de cargas. Las membranas presentan un interior negativo en relación al exterior. Esta diferencia de carga recibe el nombre de potencial de membrana. El valor del potencial de membrana en reposo es de -70 milivoltios. El milivoltio es una unidad de potencial eléctrico. El signo se coloca convencionalmente, teniendo en cuenta las condiciones del medio intracelular; el potencial lleva signo negativo porque en el medio intracelular predominan las cargas negativas.
En las membranas existen canales iónicos no regulados para el Na+ y el K+. A través de ellos, cada ión escapa siguiendo su gradiente. El Na+ ingresa a la célula y el K+, para el cual la membrana es mucho más permeable, va hacia el líquido intersticial. Sin embargo, las concentraciones iónicas se mantienen desiguales, pues la bomba de Na+ y K+ trabaja permanentemente.
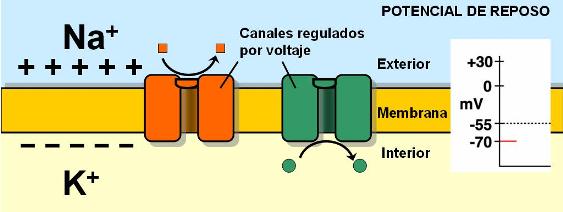 |
Cuando un estímulo físico o químico actúa sobre la membrana excitable, que mantiene un potencial de reposo, la membrana reacciona. Si el estímulo es suficientemente intenso, es decir si supera el umbral, la membrana convierte a ese estímulo en un impulso nervioso.
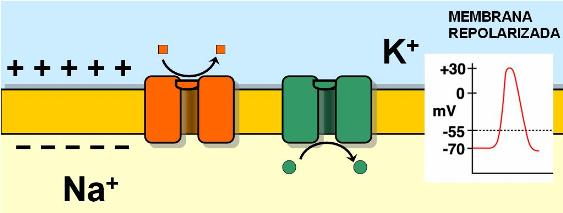
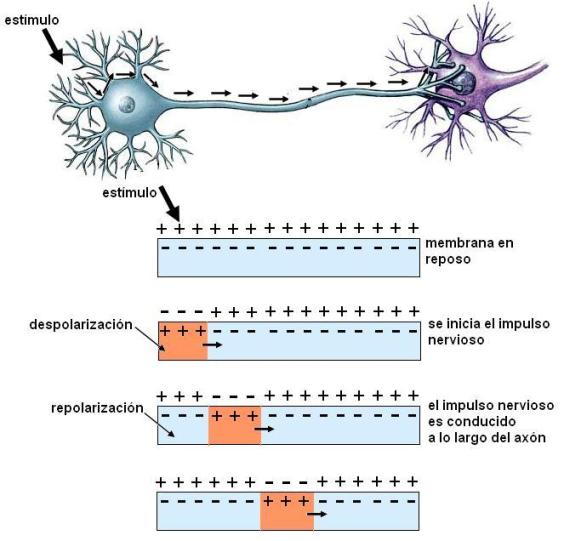
El mecanismo por el cual un estímulo desencadena el impulso nervioso consiste en una alteración de la permeabilidad de la membrana. Las membranas de las neuronas tienen canales iónicos regulados por voltaje (por el potencial eléctrico). Los estímulos abren los canales de Na+ regulados por voltaje. La apertura de estos canales es muy rápida; cuando se abren, el Na+ ingresa a la célula en forma masiva. A medida que ingresa el Na+, el interior celular se hace cada vez más positivo. Esta inversión del potencial de reposo recibe el nombre de despolarización o potencial de acción.
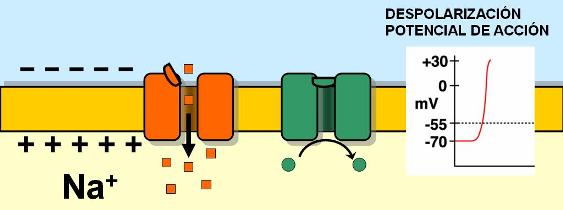 |
La despolarización en el sector de la membrana donde actuó el estímulo provoca la despolarización de los sectores vecinos. El nuevo potencial positivo en la cara interna de la membrana gatilla la apertura de los canales de sodio regulados por voltaje de la zona adyacente. Secuencialmente se abren más canales y el Na+ sigue ingresando. Así se autopropaga el potencial de acción. El potencial de acción autopropagado es lo que llamamos impulso nervioso.
El papel de la membrana en la conducción del impulso nervioso no es pasivo como el de un cable por donde pasa la corriente eléctrica. Por el contrario, la onda de despolarización se compara con el fenómeno que se produce al encender el extremo de un reguero de pólvora. Al encenderse las primeras partículas, éstas producen la combustión de las siguientes y la llama se mueve sin parar hasta el final del camino.
Los canales de Na+ regulados por voltaje se abren muy rápidamente pero permanecen abiertos por poco tiempo. Cuando el potencial llega a +35 milivoltios, los canales de Na+ cierran sus compuertas y el flujo de sodio hacia el interior de la célula se interrumpe.
Al mismo tiempo que los canales de Na+ se cierran, se abren más canales de K+ con compuertas de voltaje. Éstos tienen una apertura más lenta y prolongada que los canales de Na+. Al abrirse estos canales, el K+ sale de la célula. La pérdida de cargas positivas a través de los canales de K+ provoca que el interior de la célula se torne nuevamente negativo. Así, el potencial de la membrana retorna al valor de reposo. La recuperación del potencial de reposo negativo recibe el nombre de repolarización. Los canales de K+ se cierran cuando la repolarización se completa.
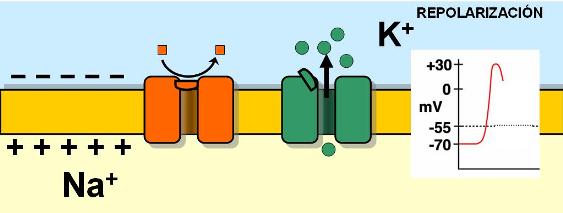 |
Durante el potencial de acción hay una primera fase en la que se produce el ingreso de Na+ y una segunda fase en la cual el ingreso de Na+ se detiene y ocurre la salida de K+, haciendo que la membrana se repolarice. Entonces, cuando se alcanza nuevamente el potencial de reposo, las concentraciones iónicas quedan invertidas, con el K+ fuera de la célula y el Na+ en el interior.
La bomba de Na+ y K+ restablece los gradientes iniciales, introduciendo nuevamente el K+ y extrayendo el Na+ de la célula. El 70% del ATP de una neurona es consumido en el trabajo de la bomba de Na+ y K+.
Todo el proceso de despolarización y repolarización de un sector de la membrana puede acontecer en menos de 1 milisegundo (mseg). A medida que el potencial de acción avanza, la parte de la membrana que queda por detrás se repolariza.
Mientras dura el potencial de acción, la neurona se halla en un período refractario absoluto, en el cual no responde a ningún estímulo. A éste le sigue un período refractario relativo, de varios milisegundos, durante el cual la neurona puede responder, pero con un umbral más alto. El disparo de un nuevo potencial de acción requiere el restablecimiento completo del estado de reposo.
Las neuronas se comportan según la ley del todo o nada. Si un estímulo alcanza el umbral, se inicia el potencial de acción y éste tiene siempre la misma intensidad. Si el estímulo no alcanza el umbral necesario, el potencial de acción no se inicia.
La diferente intensidad de nuestras sensaciones no depende de la intensidad del impulso, sino del número de neuronas estimuladas.
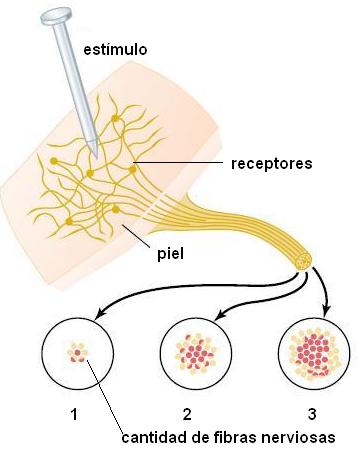 |
La cantidad de fibras nerviosas que conducen un estímulo sensitivo depende de la cantidad de receptores presentes en el área donde se aplica el estímulo. Por ejemplo, en el esquema, se muestran las áreas 1, 2 y 3, con densidad de receptores creciente. Cuantos más receptores capten el estímulo, más fibras conducirán el impulso hasta un centro nervioso y más intensa será la sensación. Las zonas más sensibles de la piel (como las yemas de los dedos) son aquéllas que poseen un mayor número de receptores por unidad de área. |
Conducción continua y conducción saltatoria Regresar
En las fibras que carecen de vaina de mielina (amielínicas) la conducción del impulso nervioso es continua. En las fibras mielínicas, en cambio, la conducción es saltatoria. En estas fibras, la vaina de mielina actúa como aislante, impidiendo el intercambio de iones a través de la membrana del axón. Las únicas zonas que pueden despolarizarse son los nódulos de Ranvier, donde la vaina de mielina se interrumpe. El impulso nervioso se propaga entonces “saltando” desde un nudo de Ranvier a otro. Esto hace que el impulso se propague más rápidamente, y también con menor gasto energético, pues requiere la despolarización y repolarización de pequeñas partes de la membrana. La velocidad de conducción varía desde 0,25m/seg en las fibras amielínicas más lentas hasta 100m/seg en las fibras mielínicas más rápidas.
Sinapsis Regresar
Las señales nerviosas se transmiten de una neurona a otra a través de una forma de comunicación intercelular llamada sinapsis. La neurona que transmite el mensaje es la presináptica y la que lo recibe, la postsináptica.
Según la forma en que se establece la comunicación, las sinapsis se clasifican en dos tipos: eléctricas y químicas.
Las sinapsis eléctricas son comunes en los invertebrados. En el hombre, se encuentran en algunas partes del SNC. Las sinapsis eléctricas consisten en el acoplamiento de las células por medio de uniones tipo nexus. A través de los conexones, el potencial de acción se propaga directamente de una célula a la otra.
La mayoría de las sinapsis en nuestro organismo son sinapsis químicas.
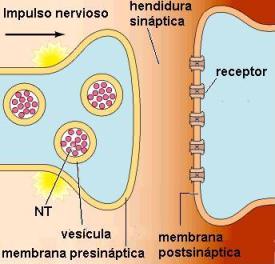
En una sinapsis química no hay contacto directo entre las células que se comunican. Las membranas de las dos neuronas están separadas por un breve espacio, la hendidura sináptica y la comunicación está mediada por una sustancia química, el neurotransmisor (NT). Las sinapsis más frecuentes son las que se producen entre el axón de una neurona y las dendritas de otra.
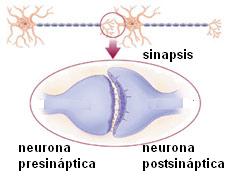 |
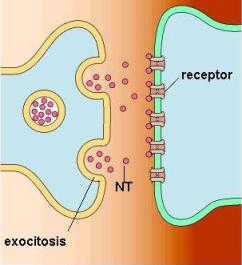
En los botones sinápticos se almacenan las vesículas que contienen los neurotransmisores. Cuando el impulso nervioso llega al terminal axónico de la neurona presináptica, las vesículas sinápticas se fusionan con la membrana plasmática. De esta forma, mediante exocitosis, los neurotransmisores son volcados al espacio sináptico.
Una vez producida la exocitosis, las membranas vesiculares se endocitan nuevamente para su reciclaje.
La exocitosis de las vesículas sinápticas es disparada por un aumento en la concentración del Ca2+ citoplasmático. Éste ingresa a los botones terminales a través de canales de calcio regulados por voltaje, que se abren con la llegada del potencial de acción. Luego, el calcio es secuestrado rápidamente dentro del botón sináptico.
Los neurotransmisores liberados en la hendidura sináptica difunden hasta la membrana postsináptica. Allí se encuentran los receptores apropiados, proteínas de membrana a las cuales se acoplan las moléculas del neurotransmisor.
 |
 |
Los receptores de los neurotransmisores pueden ser ionotrópicos o metabotrópicos.
Un receptor ionotrópico es un canal iónico regulado por ligando (se denomina ligando a una molécula que puede unirse específicamente a una proteína; en este caso el ligando es el neurotransmisor). Cuando el neurotransmisor se une a un sitio específico del receptor, éste cambia su conformación y abre su compuerta, dejando ingresar a una determinada especie iónica, por ejemplo, Na+. El ingreso del ión modifica el potencial de membrana en la neurona postsináptica.
Los receptores metabotrópicos son proteínas acopladas a proteína G. La proteína G, situada en la membrana, se activa cuando el neurotransmisor se une al receptor. La proteína G activada interactúa con una enzima encargada de fabricar una molécula llamada “segundo mensajero”. Éste es el responsable de inducir los cambios en la célula postsináptica.
La unión del neurotransmisor al receptor de la membrana postsináptica puede tener efectos excitatorios o inhibitorios. Las sinapsis excitatorias son aquéllas en las cuales el neurotransmisor desencadena un potencial de acción en la neurona postsináptica. Por el contrario, en las sinapsis inhibitorias, la membrana postsináptica se hiperpolariza, es decir, se hace aún más negativa. Esto la aleja de la posibilidad de generar un potencial de acción.
Es importante señalar que pueden existir distintos receptores para un mismo neurotransmisor. Los cambios inducidos en la célula postsináptica dependen de la interacción entre ambos.
Los neurotransmisores tienen un efecto muy breve, pues rápidamente son inactivados por alguno de los siguientes mecanismos:
- Destrucción enzimática del neurotransmisor en la hendidura sináptica.
- Recaptación del neurotransmisor en el botón terminal.
- Captación del transmisor por células gliales.
- Difusión fuera de la hendidura.
Neurotransmisores Regresar
Los neurotransmisores pueden agruparse en cuatro tipos principales:
1) Acetil colina
2) Aminas: dopamina, noradrenalina, serotonina.
3) Aminoácidos: glutamato, GABA, glicina.
4) Péptidos: opiáceos, neuropéptido Y, somatostatina.
Arco reflejo Regresar
Los actos reflejos son las respuestas más simples producidas por el sistema nervioso. Se trata de respuestas innatas, independientes de la voluntad, que ocurren rápidamente y tienen una función adaptativa. La extensión de la pierna cuando se golpea el tendón rotuliano, la acción de cerrar los ojos ante el acercamiento de un objeto o de retirar el cuerpo frente a una agresión, son ejemplos de actos reflejos.
Las estructuras que intervienen en la producción de un acto reflejo reciben, en conjunto, el nombre de arco reflejo.
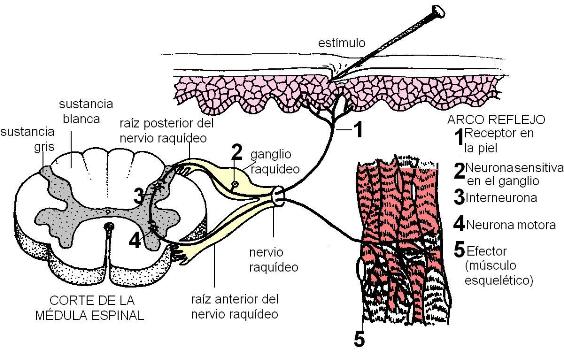
Los arcos reflejos más simples constan de un receptor, una neurona sensitiva, una neurona motora y un efector. Entre las neuronas sensitiva y motora puede interponerse una neurona de asociación.
El siguiente esquema ilustra un arco reflejo somático que se integra a nivel de la médula espinal. En este ejemplo, un estímulo doloroso actúa sobre un receptor ubicado en la piel. La información es conducida por la dendrita de una neurona sensitiva que forma parte de un nervio raquídeo. Los nervios raquídeos tienen dos raíces: una anterior y otra posterior, que luego se fusionan. Las fibras sensitivas discurren por la raíz posterior. El cuerpo de la neurona sensitiva se ubica fuera del SNC, en un ganglio raquídeo, anexo a la raíz posterior del nervio. Desde allí parte el axón de la neurona sensitiva, que a través de la raíz dorsal, ingresa a la médula espinal. En la sustancia gris de la médula, la neurona sensitiva hace sinapsis con una interneurona y ésta, con una neurona motora. El axón de la neurona motora emerge por la raíz anterior del nervio raquídeo y conduce el impulso nervioso hasta un músculo esquelético. El músculo esquelético es el efector. Cuando se produce la sinapsis entre la neurona motora y el músculo, éste ejecuta la respuesta, es decir, se contrae. La contracción del músculo genera un movimiento: el acto reflejo de retirar la parte del cuerpo que está siendo agredida por el estímulo.
 |
Niveles de funcionamiento en el SNC Regresar
El sistema nervioso de los seres humanos funciona en tres niveles, cada uno de los cuales tiene atributos funcionales específicos. Los niveles son:
Nivel medular. La médula espinal actúa como centro de reflejos, tanto somáticos como viscerales. De esta forma controla:
1) Los movimientos de la marcha.
2) Los reflejos de retirada de una parte del cuerpo ante estímulos dolorosos.
3) Los reflejos de contracción forzada en las extremidades inferiores para sostener el cuerpo en contra de la gravedad.
4) Los reflejos que controlan localmente los vasos sanguíneos, las contracciones intestinales y otras funciones viscerales.
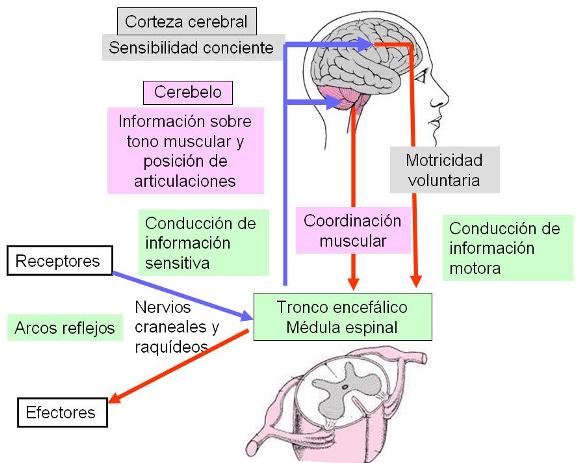
La médula también conduce información sensitiva hacia centros superiores e información motora provenientes de ellos, a través de las vías que transcurren por la sustancia blanca medular.
Por ejemplo, al producirse un acto reflejo como el descrito previamente, la información sensitiva viaja en forma ascendente hacia centros ubicados en el cerebro y en el cerebelo. La llegada de la información a centros sensitivos del cerebro es lo que produce la conciencia de la situación. En el cerebro se integra esta información y en un centro motor se origina una respuesta que refuerza el acto reflejo, como podría ser recoger el clavo. Esta respuesta es conducida a través de la médula hasta el nivel de donde emerge el nervio raquídeo, por el cual la orden llegará al músculo efector correspondiente.
La información que llega al cerebelo está relacionada con el tono muscular de los músculos implicados en el movimiento. Esta información no es conciente, pero es indispensable para la coordinación muscular. Por ejemplo, cuando el cerebro ordena recoger el clavo, ésa es una orden motora para la contracción de ciertos músculos, supongamos los flexores del antebrazo. Para que este movimiento se lleve a cabo adecuadamente, los músculos antagonistas (los extensores) deben relajarse. Controles de este tipo están a cargo del cerebelo. Las órdenes inconcientes provenientes del cerebelo también viajan por el interior de la médula hasta alcanzar el nivel del nervio raquídeo correspondiente.
 |
Nivel encefálico inferior. Este nivel comprende al cerebelo, al tronco encefálico (bulbo, protuberancia y mesencéfalo) y a los núcleos cerebrales.
El cerebelo controla el tono muscular, la postura y el equilibrio.
En el tronco encefálico se integran los reflejos somáticos de la cabeza y reflejos viscerales, como la salivación, la respiración, la presión arterial, etc. También, al igual que la médula, conduce impulsos sensitivos y motores hacia centros superiores.
Los núcleos cerebrales gobiernan muchos modelos de conducta emocional como la ira, la excitación, o la reacción ante el placer y el dolor.
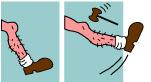
| Reflejo somático integrado en la médula espinal |
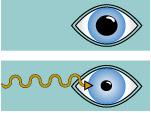
Reflejo visceral integrado en el nivel encefálico inferior |
 |
 |
| Reflejo patelar |
Reflejo pupilar |
Nivel encefálico superior. El nivel encefálico superior está representado por la corteza cerebral. En la corteza cerebral se localizan:
1) El área sensitiva somática a la cual llega la información sensitiva procedente de todo el cuerpo. Allí es cuando esta información se hace conciente.
2) Las áreas sensoriales específicas, como la auditiva o la visual.
3) El área motora somática desde donde parten las órdenes motoras para ejecutar actos voluntarios.
4) Áreas asociativas.
5) Las áreas del lenguaje.
Por último, en la corteza cerebral residen las llamadas funciones cerebrales superiores, como la memoria, el pensamiento y la conciencia.
|
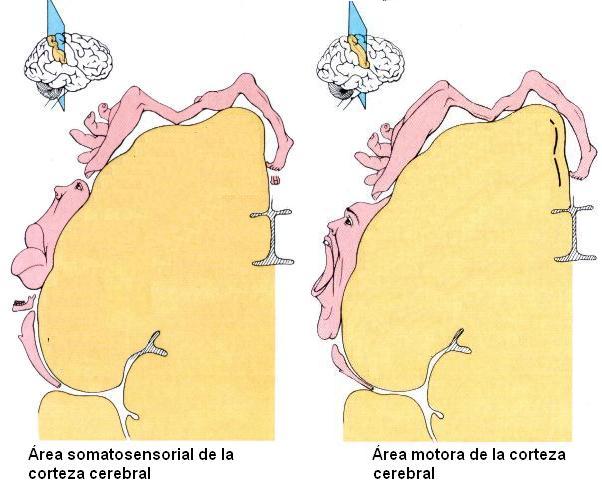 |
Los "homúnculos" indican la proporción en que las distintas partes del cuerpo se hallan representadas a nivel de las áreas sensitiva y motora de la corteza cerebral. Obsérvese la importancia de las áreas que reciben información sensitiva de los labios y los dedos, así como la extensión de las áreas motoras que controlan los músculos de la mano y la cara. |
Sistema nervioso autónomo (SNA) Regresar
El sistema nervioso autónomo o neurovegetativo ejerce el control involuntario y relativamente lento de las funciones viscerales, tales como el aporte de nutrientes, la eliminación de desechos y la distribución del riego sanguíneo.
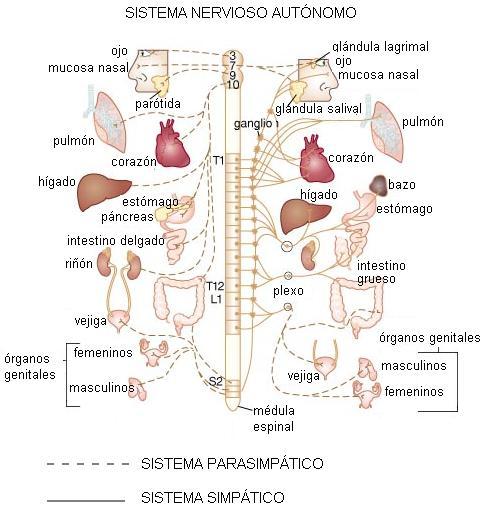
El SNA consta de dos divisiones: el sistema simpático (S) y el parasimpático (PS).
Los órganos controlados por el SNA tienen una doble inervación, pues reciben tanto nervios de la división simpática como de la parasimpática.
Las divisiones del SNA tienen funciones antagónicas, por lo cual las respuestas de los órganos efectores son la resultante de las órdenes recibidas a través de cada división.
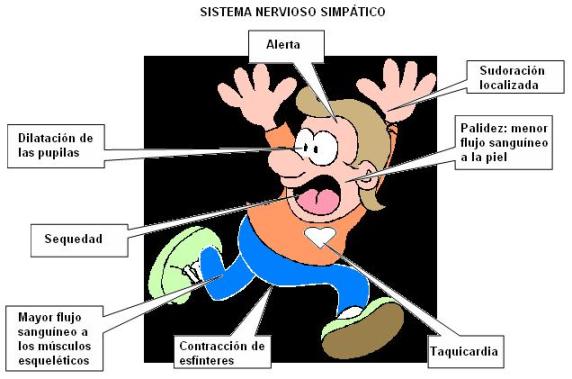 |
La función fisiológica del sistema simpático consiste en preparar al organismo para las respuestas estresantes, que podrían condensarse en la secuencia “huida, miedo o lucha”. Prepara para realizar un ejercicio físico repentino y extenuante y aumenta el estado de vigilancia o alerta. |
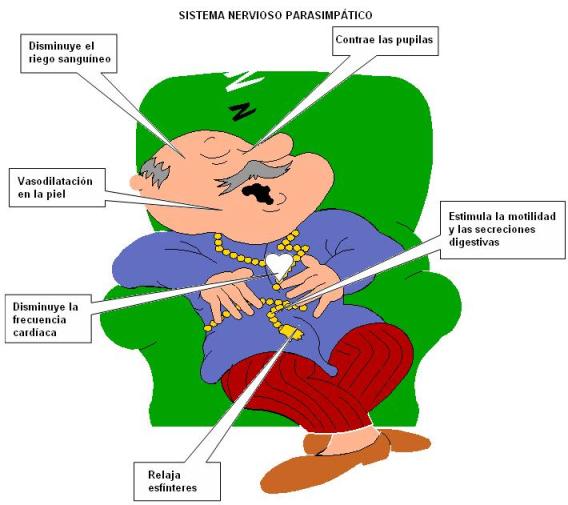 |
El sistema parasimpático ejerce acciones de control de las funciones corporales en situaciones no estresantes. Por ejemplo, existe un predominio del sistema PS durante la digestión y el reposo. En general estimula las funciones reparadoras del organismo.
|
Todos los nervios autónomos llevan fibras exclusivamente motoras. Estos nervios están constituidos por dos tipos de neuronas: las preganglionares y las posganglionares. Los cuerpos de las neuronas preganglionares están situados en el SNC. Sus axones son fibras mielinizadas. Las neuronas preganglionares hacen sinapsis con las posganglionares, cuyos cuerpos se encuentran dentro de un ganglio, en el trayecto del nervio. Los axones de las neuronas posganglionares son fibras amielínicas y hacen sinapsis con el órgano efector.
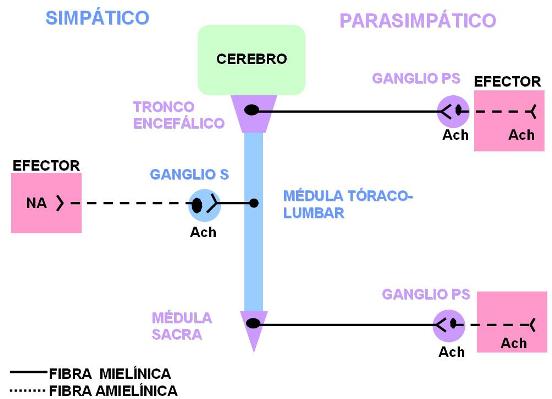 |
En el sistema simpático, el cuerpo de la neurona preganglionar se ubica en la médula espinal de las regiones torácica y lumbar. La fibra preganglionar emerge por el interior de la raíz anterior de los nervios raquídeos correspondientes a esos niveles. Luego se separa del nervio raquídeo para ingresar a un ganglio simpático prevertebral o paravertebral, donde está el cuerpo de la neurona posganglionar, con la cual hace sinapsis. El trayecto de esta fibra es corto. Desde los ganglios simpáticos sale la fibra posganglionar, más larga, que llega hasta el efector. En algunos casos, la fibra posganglionar se incorpora nuevamente al nervio raquídeo; en otros casos, forma parte de un nervio simpático.
Los cuerpos de las neuronas preganglionares del sistema parasimpático se localizan en el tronco encefálico y en la médula espinal de la región sacra. Las fibras preganglionares son largas y discurren por nervios craneales o nervios raquídeos, según el nivel donde está ubicado el cuerpo neuronal. Los ganglios parasimpáticos se encuentran en las paredes de los órganos efectores. Por lo tanto, las fibras de las neuronas posganglionares tienen cortos trayectos.
 |
|
|