|
 |
 |
 |
 |
 |
|
|
|||||
|
|||||||||||||||||||||
|
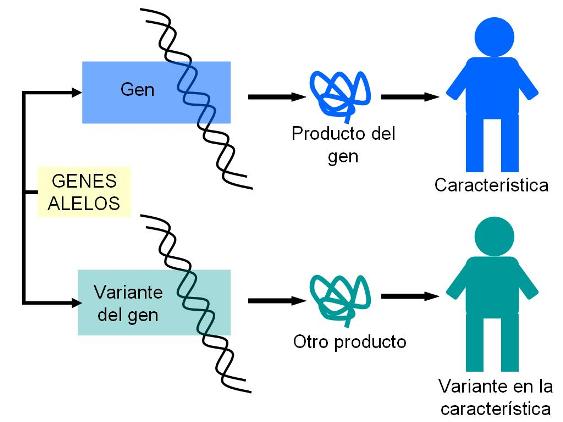
Capítulo 18: Introducción a la Genética Introducción Regresar Los genes son secuencias de ADN que codifican productos celulares (ARN, proteínas), los cuales se traducen en la aparición de un determinado rasgo o carácter.
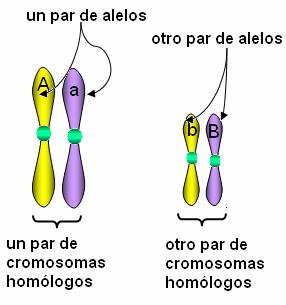
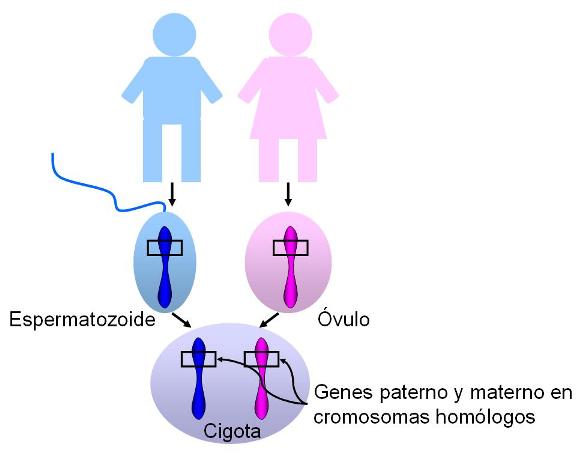
Independientemente de las variantes existentes de un gen, cada individuo sólo hereda dos copias del mismo, una de la madre y otra del padre. Dichas copias están ubicadas en el mismo locus (lugar) de los cromosomas que forman el par de homólogos.
Los alelos pueden ser dominantes o recesivos. Se denomina alelo dominante a aquél que siempre se expresa, en tanto se denomina recesivo al alelo que solo se expresa en ausencia del dominante.
El genotipo es la combinación de alelos que hereda un individuo para un rasgo determinado.
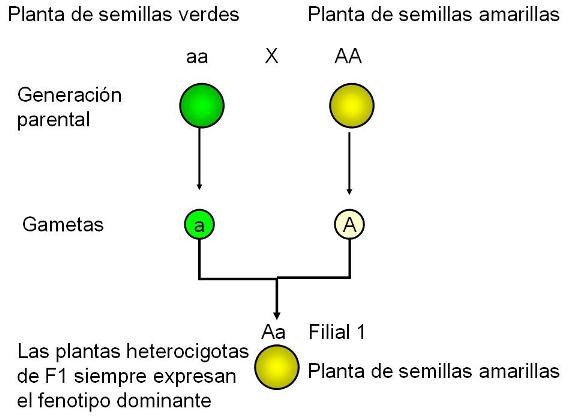
Si suponemos dos alelos de un gen, A y a, un individuo puede heredar una de las siguientes combinaciones: AA, Aa y aa. La combinación AA corresponde al genotipo homocigota dominante y la combinación aa al genotipo homocigota recesivo, en tanto la combinación Aa recibe el nombre de genotipo heterocigota.
El fenotipo es el rasgo observable que resulta de un genotipo determinado. Los individuos con genotipo homocigota dominante y heterocigota exhiben el mismo fenotipo, ya que en ambos solo se expresa el alelo dominante. El homocigota recesivo tiene un fenotipo distinto, pues es el único en el cual se manifiesta la variante recesiva del gen.
Las Leyes de Mendel Regresar
En 1868, Gregor Mendel enunció las leyes de la herencia que llevan su nombre y aún hoy están vigentes. Cuando todavía no se conocía la verdadera naturaleza química de los genes, ni su ubicación en los cromosomas, Mendel logró descubrir la forma como los genes se transmiten a la descendencia. Sus leyes surgieron como corolario de los resultados que obtuvo al analizar la herencia de algunas características de las plantas de arvejilla, que él mismo cultivaba y cruzaba para tal fin, en el monasterio agustino de Brünn, del cual llegó a ser abad. A partir de sus leyes es posible predecir los resultados de una cruza, mediante relaciones matemáticas sencillas.
La 1° Ley de Mendel: Ley de la segregación de los genes Regresar Mendel descubrió que cada individuo es portador de dos genes para cada característica, uno heredado del padre y otro de la madre, y que estos genes se segregan durante la formación de gametas. Hoy sabemos que las dos copias que heredamos de un gen se ubican en el mismo locus de dos cromosomas homólogos. Teniendo en cuenta que la formación de gametas involucra una meiosis, es claro que los dos genes que codifican la misma característica se separan durante la gametogénesis, cuando los cromosomas homólogos pasan a células hijas diferentes al concluir la primera división meiótica. Penetrancia de un gen. El ambiente y el fenotipo Regresar No siempre un genotipo expresa el fenotipo esperado. Por ejemplo, en las personas, un gen dominante (P) codifica la polidactilia, es decir la presencia de seis dedos. El alelo recesivo (p) codifica un número normal de dedos. Sin embargo, existen personas portadoras del gen dominante (P_) que no presentan polidactilia. En estos casos el gen para la polidactilia no tiene una penetrancia completa. Dominancia completa, dominancia incompleta y codominancia Regresar Se denomina dominancia completa a aquélla en la cual el individuo heterocigota presenta el mismo fenotipo que el homocigota dominante. Por ejemplo, en los cruzamientos de Mendel, la descendencia entre plantas de semillas amarillas y plantas de semillas verdes, ambas homocigotas, siempre daba como resultado plantas de semillas amarillas. El carácter “amarillo” tiene dominancia completa sobre el carácter “verde”.
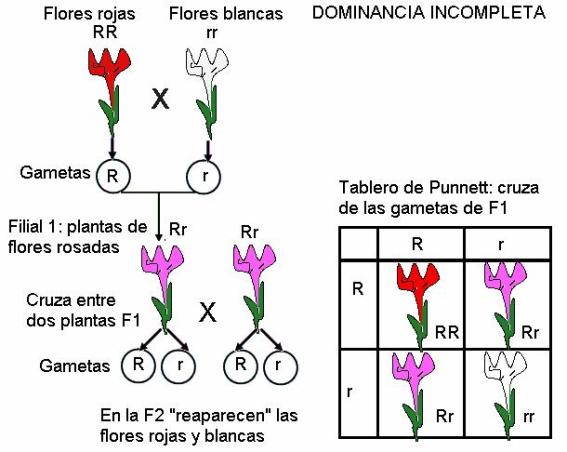
En las personas, la capacidad de doblar la lengua en “U” es debida a un gen que tiene dominancia completa sobre el alelo que determina la incapacidad de hacerlo. Así, los homocigotas dominantes y los heterocigotas para este gen, pueden enroscar la lengua; en cambio, los homocigotas recesivos son incapaces de hacerlo. En algunos casos, el gen dominante no logra encubrir por completo la expresión del gen recesivo, sino que ambos se expresan parcialmente. En los individuos heterocigotas aparece entonces un tercer fenotipo, diferente del dominante y el recesivo, e intermedio entre los dos. A estos tipos de herencia se les da el nombre de “dominancia incompleta”. Por ejemplo, en ciertas plantas, las flores pueden ser blancas, rojas o rosadas. El fenotipo “flores rosadas” se observa en los individuos heterocigotas, como resultado de la expresión parcial de los alelos que codifican los colores rojo y blanco. Cabe aclarar que, sin embargo, la aparente “mezcla” solo se produce a nivel fenotípico, ya que los alelos mantienen su individualidad, tal como lo advirtió Mendel. Así, de la cruza de dos individuos rosados, se obtendrá una progenie 25% roja, 25% blanca y 50% rosada.
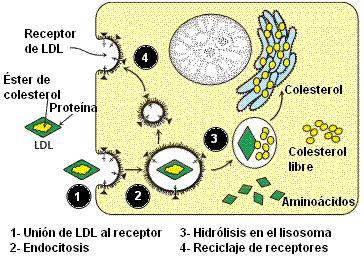
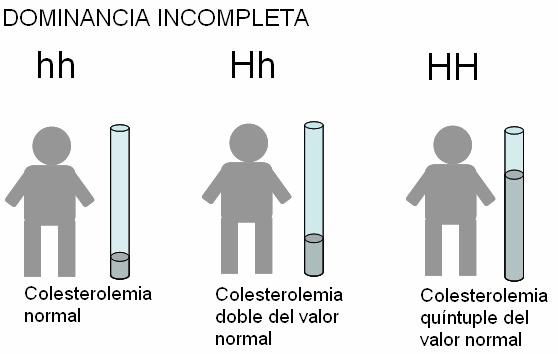
En el hombre, la hipercolesterolemia familiar es una enfermedad de origen genético, caracterizada por altos niveles de LDL en sangre, lo que predispone a la aterosclerosis y alto riesgo de enfermedad cardiovascular precoz. El trastorno se debe a la herencia de un receptor de LDL anómalo, por lo que las células no pueden endocitar estas lipoproteínas. Las LDL se acumulan en sangre y se depositan en las paredes de los vasos sanguíneos, causando su obstrucción. El alelo que codifica el receptor de LDL anormal es dominante (H) sobre el alelo que determina un receptor normal (h). Los homocigotas recesivos tienen colesterolemia normal. Las personas homocigotas dominantes (HH), tienen niveles de colesterolemia de cinco veces el valor normal, y están expuestas a padecer ataques cardíacos desde los 2 años de edad. Los heterocigotas (Hh) padecen la enfermedad, pero presentan un fenotipo intermedio, ya que su nivel de colesterol en sangre es cercano al doble del normal, y sufren ataques cardíacos alrededor de los 30 años.
Ciertos alelos son codominantes. Por ejemplo, en las personas existen tres alelos (alelos múltiples) que codifican antígenos (también llamados aglutinógenos) presentes en la membrana de los glóbulos rojos. El genotipo para este rasgo determina el grupo sanguíneo (fenotipo) que presentará el individuo. Como se puede observar en la siguiente tabla, los alelos IA e IB codifican los antígenos A y B, respectivamente, y dominan sobre el alelo i, que codifica la ausencia de antígeno. Los alelos IA e IB son codominantes. Esto resulta en la expresión de ambos antígenos en la membrana de los glóbulos rojos, en los individuos portadores de los dos alelos.
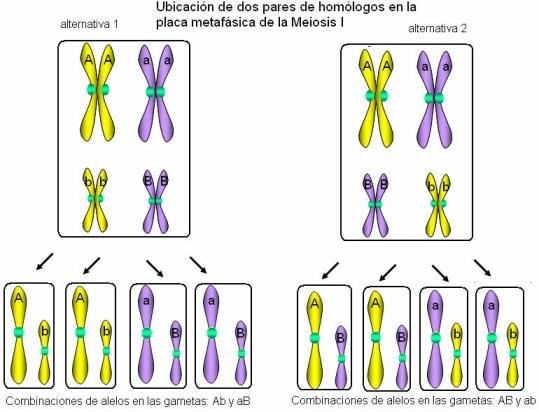
Cabe aclarar, no obstante las diferencias establecidas, que los tipos de dominancia tienen fronteras variables de acuerdo con el tipo de característica en cuestión y el nivel al cual se examine el fenotipo. La 2° Ley de Mendel: Ley de la segregación independiente Regresar La segunda ley de Mendel se refiere a la herencia simultánea de dos o más caracteres. Establece que esta herencia es independiente. Esto se verifica siempre para genes que estén ubicados en distintos pares de homólogos, ya que éstos se segregan al azar durante la gametogénesis, en la anafase I.
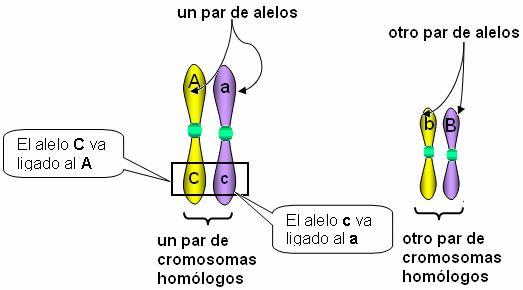
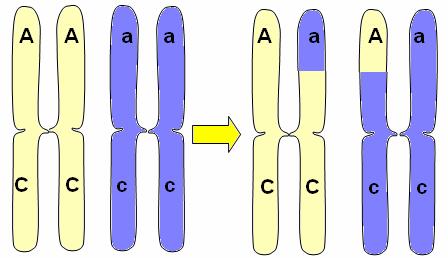
Ligamiento Regresar Se denomina ligamiento a la herencia conjunta de los genes que están ubicados en el mismo cromosoma. Dado que los cromosomas pasan completos a una de las gametas durante la meiosis, los genes de un mismo cromosoma se heredan juntos.
En el esquema, los pares de genes A/a y B/b se heredan independientemente, pues están ubicados en distintos pares de homólogos. Así, las gametas pueden llevar las combinaciones AB, Ab, aB y ab.
Herencia poligénica Regresar Si bien hasta ahora hemos analizado caracteres afectados por un solo par de genes, muchas características fenotípicas son el resultado de la interacción entre varios pares de genes.
Rasgos autosómicos, ligados al sexo y holándricos Regresar El cariotipo humano está constituido por 44 cromosomas somáticos y 2 cromosomas sexuales. El par sexual es el que determina el sexo cromosómico.
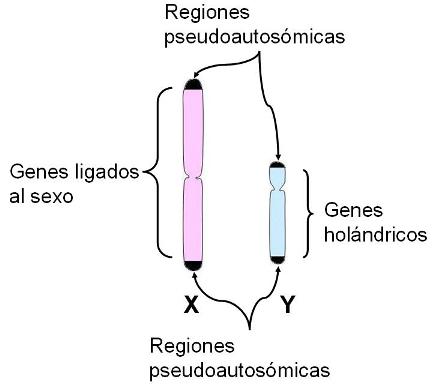
Los cromosomas sexuales Regresar El cromosoma X es submetacéntrico. La mayoría de los genes de los cromosomas sexuales se encuentran en el cromosoma X (se calcula que porta entre 1.500 y 2.500 genes). De allí que la expresión “genes ligados al sexo” se haya utilizado tradicionalmente para referirse a los genes ubicados en dicho cromosoma.
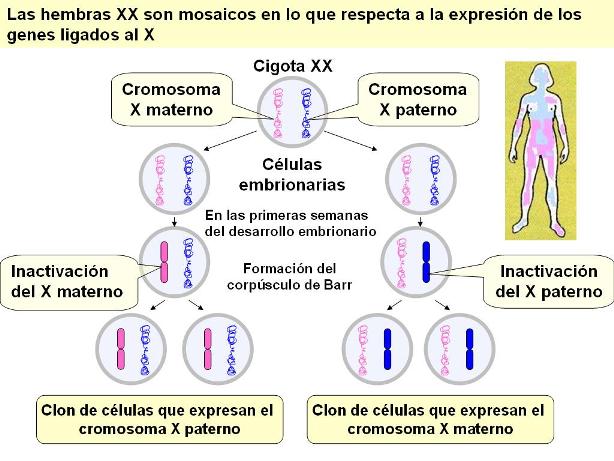
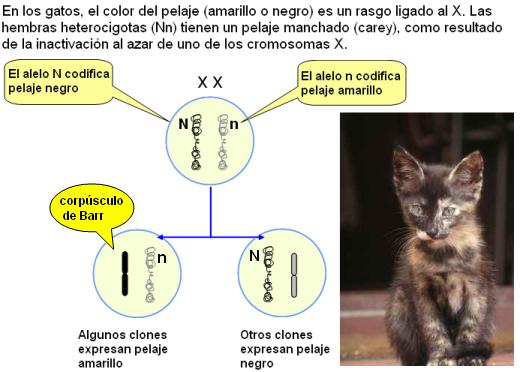
El hecho de que en el sexo XX, cada célula exprese uno solo de estos cromosomas, permite lo que se denomina “compensación de la dosis”. Esto significa que los productos codificados por los genes presentes en el X se expresan en la misma dosis que en los machos, portadores de una sola versión del cromosoma. Es decir que, funcionalmente, las hembras son hemicigóticas para los genes ligados al X. Incluso en los individuos que tienen un número anormal de cromosomas sexuales, como en el síndrome de Klinefelter (XXY) o Turner (X0) se cumple la regla según la cual se inactivan todos los X menos 1.
El cromosoma Y es un cromosoma acrocéntrico, del cual aproximadamente dos tercios corresponden a secuencias cortas repetidas, en estado heterocromático. El tercio restante consta de relativamente pocos genes. Algunos de ellos se expresan en todas las células del cuerpo, pero muchos solo lo hacen en los testículos. La herencia citoplasmática Regresar Las mujeres heredan los cromosomas X de su madre y de su padre. Los varones heredan el cromosoma X de su madre y el Y de su padre. Pero todos heredamos el cromosoma mitocondrial de nuestra madre. En las células animales, existe una herencia citoplasmática además de la nuclear: el ADN mitocondrial. Esta herencia se produce por vía exclusivamente materna, pues como ya vimos al tratar el tema fecundación, es el óvulo el que aporta el citoplasma, con sus organoides, para formar la cigota.
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|||||||||||||||


 Grupo A
Grupo A  Anti-B
Anti-B Grupo B
Grupo B  Anti-A
Anti-A Grupo AB
Grupo AB  Grupo 0
Grupo 0  Anti-A y Anti-B
Anti-A y Anti-B