|
Capítulo 16: Meiosis y gametogénesis
Reproducción sexual Regresar
La reproducción sexual es aquélla en la cual intervienen células especializadas, llamadas células sexuales o gametas. En los mamíferos, las gametas provienen de dos progenitores diferentes, masculino y femenino, y se denominan respectivamente espermatozoide y óvulo.
El espermatozoide y el óvulo se fusionan, en el proceso de fecundación, dando origen a la célula huevo o cigota, la primera célula del organismo hijo.
El descendiente reúne la información genética proveniente de ambas gametas; la mitad de su información genética es heredada del espermatozoide y la otra mitad del óvulo. La ventaja de la reproducción sexual frente a la asexual radica precisamente en la posibilidad de combinar información genética de distinta procedencia, aumentando las variantes dentro de la especie. Las especies que sólo realizan reproducción asexual transmiten a su descendencia un legado hereditario invariable, más allá de los ocasionales cambios producidos por las mutaciones.
Cada especie posee un patrón cromosómico que le es característico. El número y el tipo de cromosomas son constantes en todos los miembros de una misma especie. Se denomina cariotipo a la constitución cromosómica de un individuo.
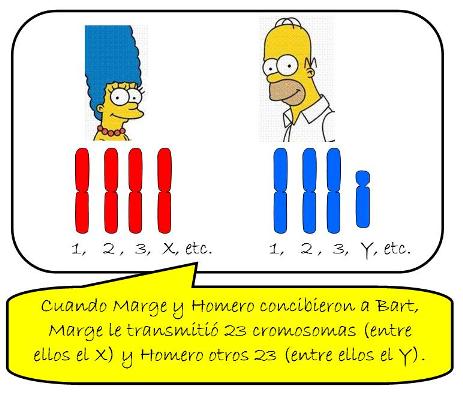
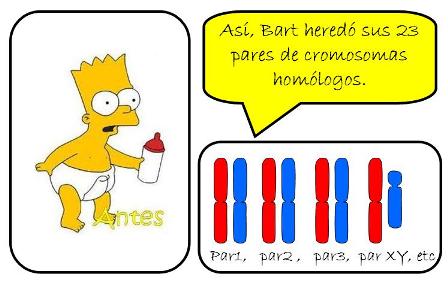
El cariotipo humano está constituido por 46 cromosomas, de los cuales 23 proceden del padre y 23 de la madre. Los 46 cromosomas no son todos distintos. Por el contrario, cada cromosoma de origen paterno tiene su contraparte de origen materno, de manera que los cromosomas pueden ser agrupados por pares. Los cromosomas que forman un par son homólogos entre sí.
El cariotipo humano, entonces, está formado por 23 pares de cromosomas homólogos. Las células cuyos cromosomas se presentan por pares reciben el nombre de diploides (diplo: doble) y se simbolizan “2n”. Nuestras células son “2n=46”.
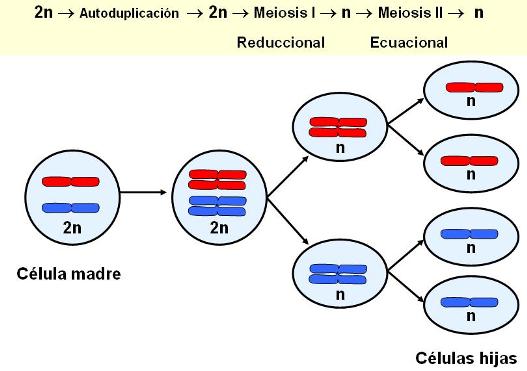
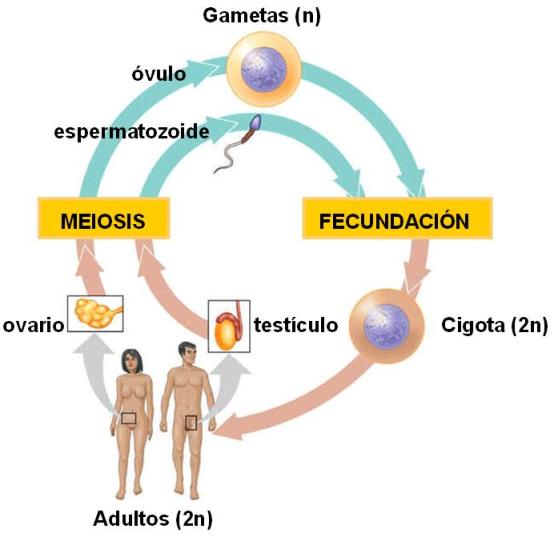
Resulta evidente que en los ciclos biológicos que incluyen reproducción sexual, en los que ocurre la fusión de gametas, debe existir algún mecanismo para evitar que el número cromosómico se duplique en cada generación. Este mecanismo es la meiosis.
La meiosis es un tipo de división celular en el cual las células hijas resultantes poseen la mitad de los cromosomas que tenía la célula original. Las células obtenidas por meiosis son haploides (haplo: simple), pues portan un juego simple de cromosomas. Ese juego simple de cromosomas está conformado por un cromosoma de cada par de homólogos.
Los animales con reproducción sexual tienen un ciclo biológico diploide. En este tipo de ciclo, los individuos son diploides y el único estadío haploide de la especie lo constituyen las gametas. La meiosis se lleva a cabo en las gónadas (ovario y testículo) con el fin de obtener gametas haploides a partir de células madre diploides.
Las gametas humanas tienen una dotación cromosómica “n=23”, lo que significa que son haploides (n) y poseen un solo cromosoma de cada par.
Al producirse la fecundación, la cigota reconstituye el número diploide. Dado que el organismo multicelular deriva de sucesivas divisiones mitóticas de la cigota, todas las células que forman a un individuo son diploides y genéticamente idénticas a aquélla.
 |
Cariotipo humano Regresar
El cariotipo humano consta de 23 pares de cromosomas homólogos. Cada par de homólogos posee un representante de origen materno y otro de origen paterno.
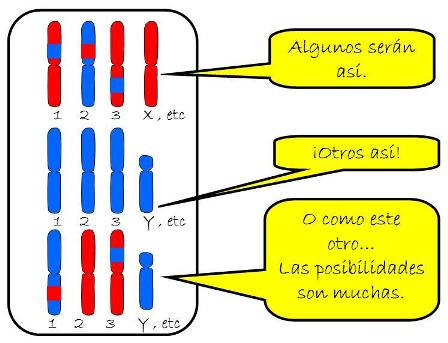
Aunque los cromosomas que forman un par de homólogos tienen la misma forma y tamaño, no son idénticos. Los cromosomas homólogos llevan genes que codifican los mismos caracteres, pero es posible que porten versiones distintas de estos genes.
Cada versión posible de un gen se denomina alelo. Por ejemplo, el gen que codifica el color de ojos presenta varios alelos: marrón, verde, azul, gris. Un individuo puede heredar simultáneamente un cromosoma con el alelo para color de ojos que codifica “gris” y el homólogo con el alelo que codifica ”marrón”, ya que los dos cromosomas homólogos provienen de individuos distintos (el padre y la madre).
Así, los individuos diploides portan un doble juego de genes al sumar los 23 homólogos paternos más los 23 maternos. En ese doble juego, cada par de genes podrá estar formado por alelos iguales o distintos.
El cariotipo de un individuo se representa mediante un gráfico en el cual los cromosomas se colocan ordenados por forma y tamaño y agrupados en pares de homólogos.
Para representar un cariotipo se fotografían los cromosomas en el momento en que son visibles al microscopio, es decir, durante la mitosis. Generalmente los cromosomas son fotografiados en la metafase mitótica, pues es en esta etapa cuando alcanzan el mayor grado de condensación y pueden observarse mejor. Por este motivo los cromosomas se ven duplicados, ya que en metafase cada uno aún conserva las dos cromátides hermanas. Una vez tomada la fotografía, las imágenes de los cromosomas se recortan y ordenan.
El siguiente esquema es una representación gráfica del cariotipo humano, tomado de una célula somática (corporal) diploide durante la metafase mitótica. En él se observan los 23 pares de homólogos que lo conforman.
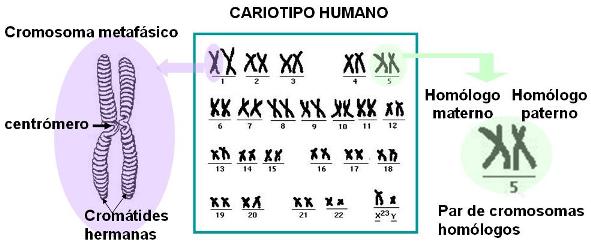 |
Los 23 pares de cromosomas homólogos presentes en cualquiera de las células somáticas humanas se clasifican en dos tipos:
• Cromosomas somáticos o autosomas: son los cromosomas de los pares 1 al 22. Los genes portados por estos cromosomas codifican rasgos somáticos, comunes a ambos sexos.
• Cromosomas sexuales, gonosomas o alosomas: son los cromosomas del par 23. En estos cromosomas se ubican los genes que determinan las diferencias entre el sexo masculino y el femenino. El par de cromosomas sexuales puede estar constituido por dos cromosomas “X” (XX) o bien por un cromosoma “X” y otro “Y” (XY). Las células de las mujeres llevan el par XX y las de los varones, el par XY.
Por lo tanto, la dotación cromosómica de una célula somática nucleada de cualquier tejido, perteneciente a una mujer, será la siguiente:
| CÉLULA SOMÁTICA FEMENINA = 2n = 46 = 22 pares de autosomas + XX |
En tanto, la dotación cromosómica de una célula somática nucleada de cualquier tejido, perteneciente a un varón, será:
| CÉLULA SOMÁTICA MASCULINA = 2n = 46 = 22 pares de autosomas + XY |
Las gametas llevan solamente 23 cromosomas. La meiosis (división celular que les da origen a partir de células especializadas del ovario y el testículo) posibilita la separación de los cromosomas homólogos, de manera que cada gameta reciba un cromosoma de cada par. Esta separación afecta tanto a los cromosomas somáticos como a los sexuales.
La dotación cromosómica de las gametas es:
| GAMETA = n = 23 = 22 autosomas + 1 cromosoma sexual |
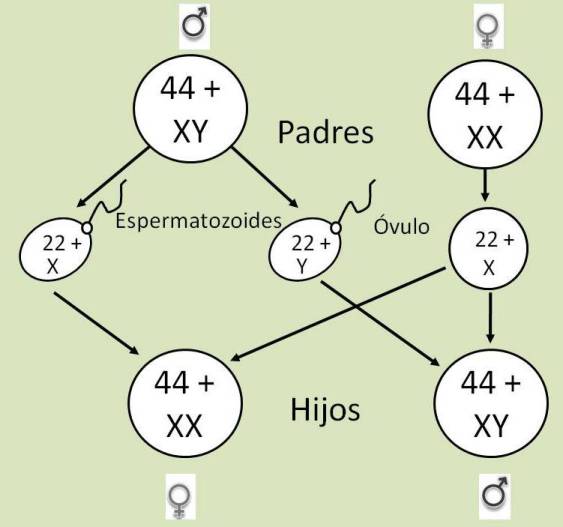
El cromosoma sexual presente en las gametas femeninas es siempre un cromosoma X. Pero en las gametas masculinas, el cromosoma sexual puede ser el X o el Y. La mitad de los espermatozoides que produce un varón lleva el cromosoma X y la otra mitad, el Y.
| ÓVULO = n = 23 = 22 autosomas + X |
| ESPERMATOZOIDE = n = 23 = 22 autosomas + X |
| ESPERMATOZOIDE = n = 23 = 22 autosomas + Y |
Determinación cromosómica del sexo Regresar
En los mamíferos, el sexo cromosómico queda determinado en el momento de la fecundación, de acuerdo con los cromosomas que hereda la cigota.
Si un óvulo es fecundado por un espermatozoide portador del cromosoma X, la cigota resulta femenina.
Si, en cambio, el óvulo es fecundado por un espermatozoide portador del cromosoma Y, la cigota resultante es de sexo masculino.
Meiosis Regresar
La meiosis es un tipo especial de división celular. Las células que inician la meiosis son siempre diploides. En los animales, sólo las células de la línea germinal, ubicadas en las gónadas, realizan la meiosis. El producto de la misma son las gametas.
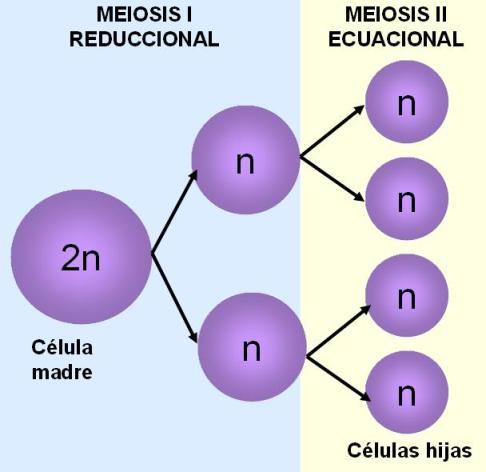
La meiosis consiste en dos divisiones sucesivas, designadas convencionalmente meiosis I y meiosis II. Durante la interfase que precede a la meiosis, los cromosomas se duplican. Al terminar la meiosis I se obtienen dos células hijas, ambas haploides. La meiosis I es reduccional, pues produce células haploides a partir de una célula diploide.
Las células hijas pueden pasar por un período de intercinesis, pero esta vez, el material cromosómico no se duplica. A continuación, cada célula hija inicia la meiosis II. El resultado final son cuatro células hijas haploides. La meiosis II es ecuacional, ya que comienza y termina con células haploides.
 |
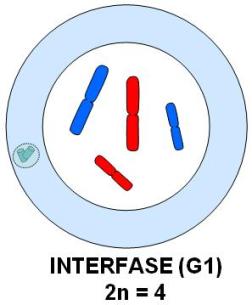
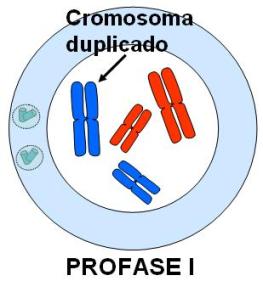
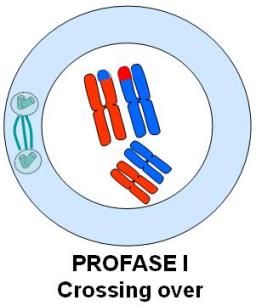
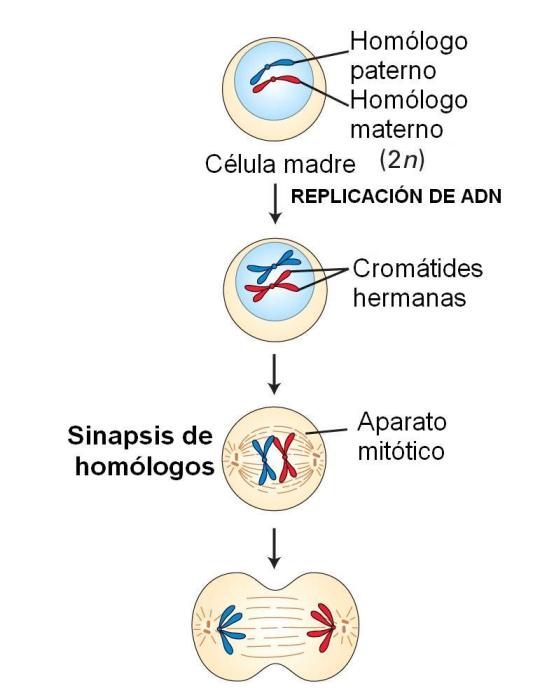
En cada división meiótica se reconocen las mismas etapas que en una mitosis: profase, metafase, anafase y telofase. Los esquemas que acompañan la explicación ilustran cada una de estas etapas en una célula hipotética 2n=4. El origen materno o paterno de los cromosomas se identifica con distinto color (rojo y azul). Profase I
Se inicia con los cromosomas duplicados, que comienzan su condensación.
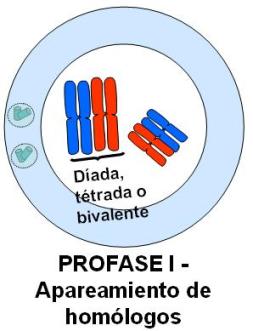
En esta etapa, los cromosomas homólogos se aparean. El apareamiento o sinapsis de los homólogos es mediado por un complejo proteico, el complejo sinaptonémico. La sinapsis da lugar a la formación de grupos llamados tétradas, bivalentes o díadas, pues cada uno de ellos consta de un par de cromosomas homólogos duplicados, es decir, de 4 cromátides.
Una vez apareados, entre los homólogos se produce el crossing over o entrecruzamiento genético. Este fenómeno consiste en un intercambio de fragmentos equivalentes, en uno o más puntos, entre cromátidas no hermanas. Los puntos de entrecruzamiento reciben el nombre de quiasmas. Como resultado se obtienen cromátides con información recombinada de origen materno y paterno. Si bien el esquema lo representa en un solo par de homólogos, el crossing over acontece en cada uno de los pares cromosómicos.
Entre tanto, en el citoplasma se forma el huso, que se va extendiendo hacia los polos. La envoltura nuclear comienza su disgregación. Cuando ésta se halla completamente disgregada, se considera concluida la profase.
Metafase I
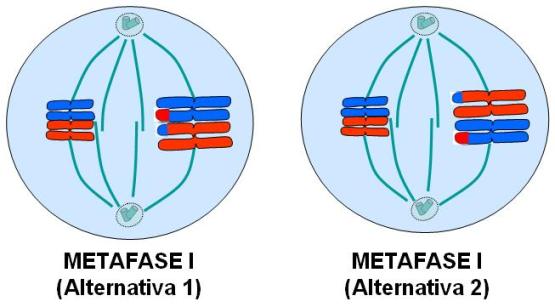
Los cromosomas homólogos se ubican en el plano ecuatorial. Los dos homólogos de un par se unen a la misma fibra del huso. Significa que durante la anafase I se separarán cromosomas homólogos.
La ubicación de los homólogos en el huso es azarosa. Esto es: cualquier homólogo de un par, materno o paterno (en el gráfico diferenciados por los colores rojo y azul), puede orientarse indistintamente hacia un polo u otro. Por lo tanto, para cada par de homólogos hay dos posibilidades, igualmente probables.
Como los homólogos de un par no llevan idéntica información genética, las células hijas resultantes heredarán diferentes caracteres, de acuerdo con la forma en que los cromosomas se dispongan en la placa ecuatorial. Esto se ilustra en el esquema, donde se muestran las dos alternativas posibles para la célula hipotética 2n=4 de nuestro ejemplo. Si la meiosis procediera según la alternativa 1, se obtendrían dos células hijas con combinaciones cromosómicas diferentes entre sí y respecto de las que resultarían al seguir la alternativa 2.
 |
Anafase I
En esta etapa tiene lugar la separación al azar de los homólogos, que migran hacia los polos opuestos de la célula.
La separación al azar de los homólogos genera diferentes combinaciones cromosómicas en las células hijas, por lo ya expuesto en el ítem anterior.
Existen dos alternativas por cada par de homólogos. Se calcula entonces, que el número de células hijas con diferentes combinaciones cromosómicas es de 2n, donde n = número de pares homólogos (número haploide).
En el ejemplo ilustrado, habría 4 combinaciones (2 por cada alternativa).
Para la célula humana, 2n es igual a 223, lo que equivale a ¡8.388.308 variantes!
La separación al azar de los homólogos en la anafase I genera una gran diversidad genética.
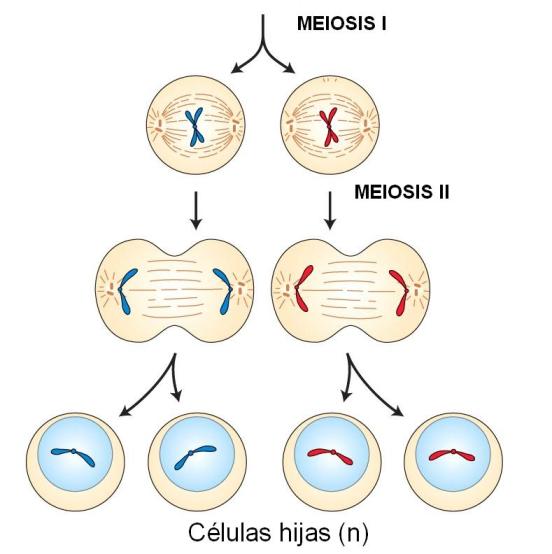
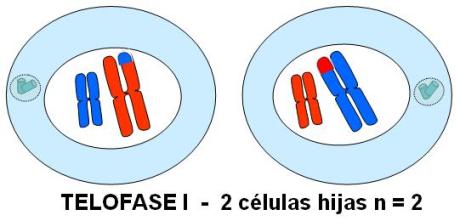
Telofase I
Los homólogos llegan a polos opuestos de la célula. En cada polo se reorganiza la envoltura nuclear alrededor de los cromosomas, formándose dos núcleos hijos. Cada núcleo hijo contiene un cromosoma de cada par de homólogos. Por lo tanto, se trata de núcleos haploides,
a pesar de que cada cromosoma aún es doble.
Los cromosomas se descondensan, volviendo a adquirir el aspecto interfásico.
El huso se desorganiza. La citocinesis puede ocurrir en esta etapa o diferirse hasta el término de la meiosis II. Si se produce la citocinesis, se obtienen dos células hijas.
En el esquema se representan las células hijas correspondientes a la alternativa 2.
 |
Profase II
Cada célula hija obtenida de la meiosis I inicia la meiosis II, sin previa duplicación del material genético. Sin embargo, cada cromosoma consta de dos cromátides hermanas, debido a la duplicación del ADN ocurrida antes de comenzar la meiosis I.
La profase II es similar a una profase mitótica: los cromosomas se condensan, la envoltura nuclear se disgrega y se forma el huso. Este último adopta una dirección perpendicular con respecto al de la meiosis I.
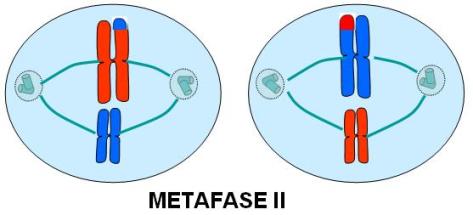
Metafase II
Los cromosomas se ubican en el plano ecuatorial. Cada cromosoma, con sus dos cromátides hermanas, se une independientemente a una fibra del huso.
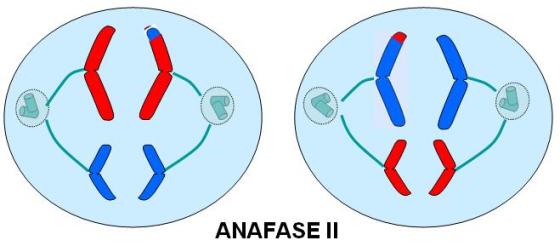
Anafase II
Las cromátides hermanas de cada cromosoma se separan al azar, migrando hacia polos opuestos.
Obsérvese que, a causa del crossing over, las cromátides hermanas ya no son idénticas como lo eran al iniciarse la profase I. Como ambas cromátides pueden dirigirse indistintamente a cualquiera de los dos polos, para cada cromosoma hay dos alternativas posibles. Combinando las distintas alternativas de todos los cromosomas, se amplía enormemente el número de variantes potenciales en las células hijas.
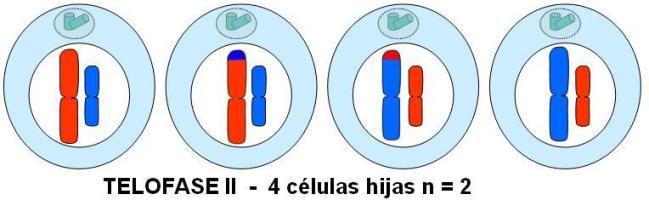
Telofase II
Las cromátides hermanas, ahora cromosomas hijos, llegan a los polos. En cada polo, los cromosomas hijos son rodeados por una envoltura nuclear y se descondensan.
Se desorganiza el huso y se completa la división citoplasmática.
El resultado final son 4 células hijas haploides. La combinación de cromosomas y la combinación de genes en cada cromosoma es diferente para las cuatro células hijas.
En conclusión, la meiosis posibilita:
| 1. La obtención de células haploides, ya que después de duplicar el material genético una sola vez, se realizan dos divisiones sucesivas. En la primera de ellas se separan los cromosomas homólogos; en la segunda, se separan las cromátides hermanas. |
| 2. La generación de diversidad dentro de la especie, puesto que las células hijas son distintas entre sí, y probablemente lo sean en cada meiosis que realiza un individuo. Esto se debe a tres fenómenos: el crossing over, la separación al azar de los homólogos y la separación al azar de la cromátides hermanas. |
Gametogénesis Regresar
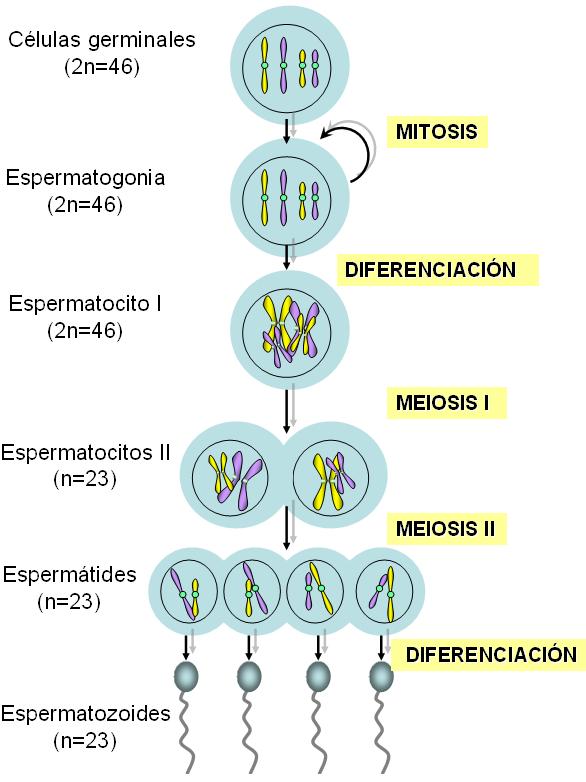
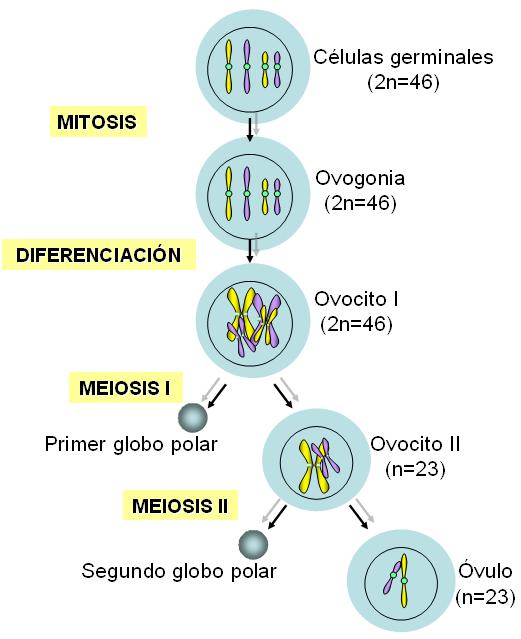
La producción de gametas mediante la división meiótica recibe el nombre de gametogénesis. La gametogénesis tiene lugar en las gónadas u órganos sexuales, a partir de las células germinales diploides (2n=46). Se denomina espermatogénesis a la formación de espermatozoides y ovogénesis a la formación de óvulos.
Las células germinales masculinas, o espermatogonias, realizan dos funciones: mantienen la población de células germinales y dan origen a los espermatozoides. La mantención de la población de espermatogonias se consigue mediante divisiones mitóticas, que generan más espermatogonias diploides.
La espermatogénesis se inicia en la pubertad, cuando algunas espermatogonias comienzan diferenciarse a espermatocitos primarios. Éstos experimentan la meiosis I, originando dos espermatocitos secundarios haploides (n=23). Cada espermatocito secundario completa la meiosis II, obteniéndose cuatro espermátidas, también haploides (n=23). La citocinesis tiene lugar al final del proceso.
Las espermátides ya no se dividen; solamente sufren un proceso de diferenciación, llamado espermiogénesis, para convertirse en espermatozoides. El espermatozoide ya diferenciado posee una cabeza, un cuello y una cola.
La espermiogénesis implica una pérdida de citoplasma, la formación del acrosoma, un derivado lisosómico cuyas enzimas participarán en la fecundación, y la adquisición del flagelo, que constituirá la cola del espermatozoide. Sin embargo, los espermatozoides no adquieren movilidad hasta después de su almacenamiento en el epidídimo.
Todo el proceso de espermatogénesis dura alrededor de 64 días y depende de la hormona masculina testosterona. Debido a la mantención de una población de espermatogonias, la espermatogénesis puede proseguir hasta una edad avanzada.
 |
Las células germinales femeninas, ovogonias, se reproducen por mitosis en la gónada embrionaria, hasta que, entre el tercero y el séptimo mes de la vida prenatal, se diferencian a ovocitos I. El número de éstos es finito, pues ya no existen células madre.
En la vida prenatal los ovocitos I inician la primera meiosis, pero ésta queda interrumpida en etapa de profase antes del nacimiento. En el momento del nacimiento, una niña posee alrededor de 1.000.000 de ovocitos, todos detenidos en la profase I. Así permanecen hasta la pubertad.
A partir de la pubertad, en cada ciclo menstrual, varios folículos (las estructuras que contienen los ovocitos), comienzan a madurar. Sin embargo, habitualmente sólo uno de ellos, el folículo dominante, completa su maduración. El ovocito I del folículo dominante retoma y concluye la primera división meiótica.
El resultado de la meiosis I son dos células haploides: el ovocito II y el primer globo o cuerpo polar. La citocinesis ocurre de tal forma que el ovocito II recibe casi todo el citoplasma en detrimento del globo polar. Este último muere, en tanto el ovocito II es liberado del ovario, en el proceso llamado ovulación.
El ovocito II comienza su descenso por la trompa de Falopio, donde inicia la meiosis II. Sin embargo, esta división se completa solamente si el ovocito II es fecundado. En este caso, se forman dos células: el óvulo, la gameta funcional que inmediatamente se fusiona con el núcleo del espermatozoide, y otro pequeño globo polar, que degenera. Por lo tanto, partiendo de un ovocito I, en la mujer se obtiene una sola gameta funcional.
Cuando no hay fecundación, el ovocito II muere en la trompa de Falopio, sin terminar la segunda división meiótica.
A lo largo de la vida llegan a madurar alrededor de 400 ovocitos, desde la pubertad hasta la menopausia. Los ovocitos que maduran en los últimos ciclos pueden haber demorado alrededor de 50 años en completar la división meiótica. Esto hace que las mujeres añosas que conciben tengan mayores probabilidades de generar descendencia con anormalidades en el número cromosómico.
 |
Anomalías en el número cromosómico Regresar
Errores ocurridos durante la gametogénesis pueden ocasionar la formación de gametas con un número anormal (en más o en menos) de cromosomas, alteraciones que reciben el nombre de aneuploidías. Las cigotas resultantes no suelen ser viables cuando presentan monosomías autosómicas (existencia de un solo cromosoma del par). Sin embargo, sí lo son algunas trisomías, como el síndrome de Down, que se caracteriza por la existencia de tres cromosomas 21.
Las aneuploidías más comunes observadas en los seres humanos están relacionadas con los cromosomas sexuales, como el síndrome de Turner (mujeres XO) y el síndrome de Klinefelter (varones XXY).
|
|