|
Capítulo 17: Sistema Reproductor
Introducción Regresar
Las características biológicas correspondientes a cada sexo se instauran en tres etapas diferentes a lo largo de la vida.
- En primer lugar se establece el sexo cromosómico, cuando en la fecundación se forman las cigotas XX (femenina) o XY (masculina).
- En segundo lugar, durante el desarrollo embrionario, se adquieren los caracteres sexuales primarios. Esto es, las primitivas estructuras reproductoras, que hasta la 6ª semana del desarrollo son prácticamente idénticas en varones y mujeres, comienzan su diferenciación, dando lugar a la formación de testículos u ovarios y sus respectivos anexos.
- El tercer hito se ubica en la pubertad. En esta etapa, bajo el influjo hormonal, ocurren importantes cambios, externos e internos, que conducen a la madurez sexual.
Por un lado, se produce la aparición de los caracteres sexuales secundarios. Se entiende por tales a todos aquellos caracteres que permiten diferenciar externamente al varón de la mujer adultos. Por ejemplo, la distribución del vello y de la grasa corporal, las proporciones corporales, el desarrollo de la masa muscular, el crecimiento y desarrollo de los genitales externos, el tono de voz.
Por otro lado, en la pubertad culmina el desarrollo del sistema reproductor y éste adquiere su pleno funcionamiento, marcado por el inicio de la fertilidad.
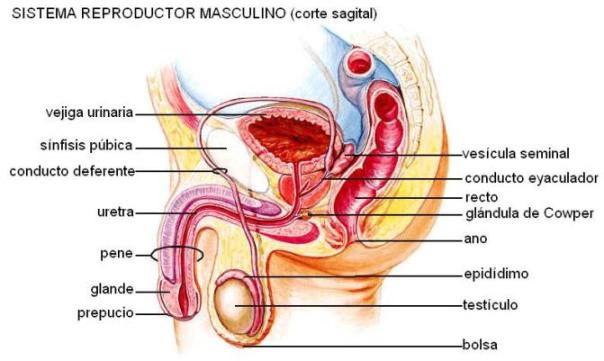
Sistema reproductor masculino Regresar
El sistema reproductor masculino consta de los siguientes órganos: • Gónadas u órganos sexuales primarios: son los testículos. Sus funciones consisten en la producción de gametas masculinas (espermatogénesis) y la secreción de hormonas sexuales masculinas.
• Conductos genitales: incluyen al epidídimo, al conducto deferente, al conducto eyaculador y a la uretra. Se encargan del transporte del semen al exterior.
• Glándulas anexas: próstata, vesículas seminales y glándulas bulbouretrales o de Cowper. Secretan los componentes del semen.
• Pene: es el órgano copulador.
Testículos Regresar
Los testículos están ubicados fuera de la cavidad abdominal, dentro de un sistema de cubiertas concéntricas que se designan en conjunto con el nombre de bolsas. Las paredes de las bolsas constan de una capa externa, la piel o escroto, de fibras musculares lisas, tejidos elástico y fibroso y una membrana dependiente del peritoneo.
Los testículos descienden hasta las bolsas desde la cavidad abdominal en el último trimestre del embarazo, atravesando las paredes del abdomen por el canal inguinal.
Su posición las bolsas permite que tengan una temperatura 2 - 3 ° C menor que la temperatura corporal interna, lo cual es necesario para la producción de los espermatozoides.
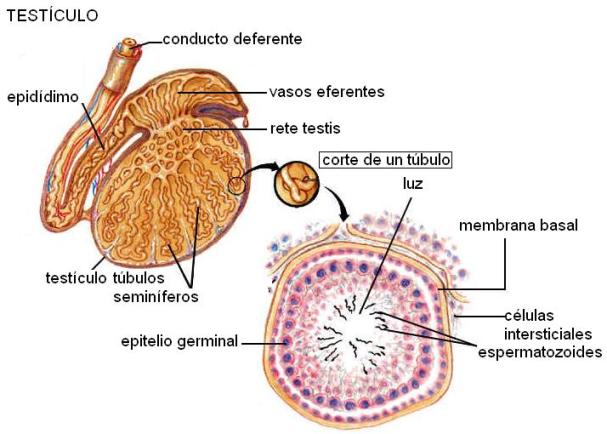 |
En el interior de los testículos se encuentran los túbulos seminíferos, donde se forman los espermatozoides a partir de las células germinales. En el tejido intersticial, ubicado entre los túbulos, se encuentran las células de Leydig, con función endócrina. Éstas secretan las hormonas esteroideas masculinas y otros mediadores locales.
Los túbulos seminíferos poseen una membrana basal, que los separa del intersticio. Las paredes de los túbulos están formadas por las células de Sertoli y las células germinales. Las células de Sertoli tienen prolongaciones que rodean y envuelven a las células germinales en desarrollo, formando la barrera hematotesticular. Además, contribuyen a la formación del líquido seminal y secretan la proteína fijadora de andrógenos y la hormona inhibina.
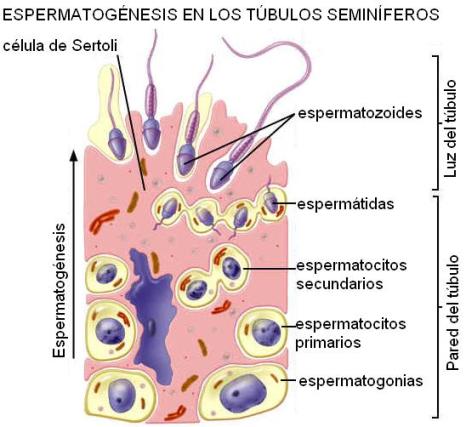 |
Las células germinales van migrando desde los bordes hacia el centro del túbulo, siempre contenidas por las células de Sertoli, a medida que avanzan en su desarrollo. Finalmente, cuando se diferencian los espermatozoides, éstos son liberados en la luz de los túbulos. Como la citocinesis no se produce sino hasta el final de la espermatogénesis, todas las células derivadas de una espermatogonia permanecen conectadas por un citoplasma común. Al culminar la diferenciación, los puentes citoplasmáticos se rompen y los espermatozoides aparecen en la luz sincrónicamente.
Los túbulos seminíferos se vacían en una red de conductillos, la rete testis, de la cual nacen los vasos eferentes. Desde allí los espermatozoides son transportados hacia el epidídimo.
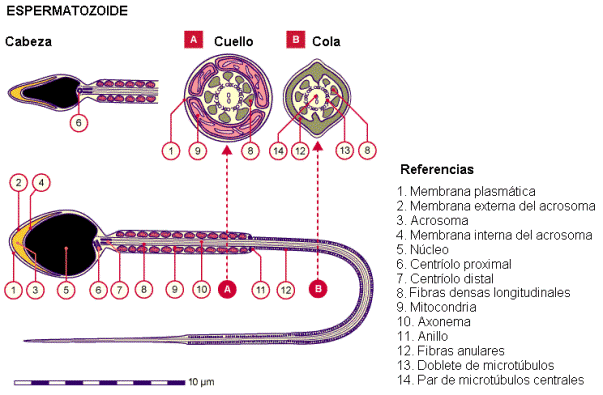
Espermiogénesis Regresar Se denomina espermiogénesis a la diferenciación de las espermátidas en espermatozoides. El proceso de transformación de espermátidas en espermatozoides incluye:
- La reducción del tamaño nuclear y la condensación del material genético.
- La formación del acrosoma. El acrosoma es una vesícula originada del aparato de Golgi, que se ubica en la cabeza del espermatozoide, a modo de casquete, envolviendo parte del núcleo. Contiene enzimas hidrolíticas que se exocitan en el momento de la fecundación, ayudando a penetrar la membrana externa del ovocito.
- El desarrollo del flagelo. Éste formará la cola del espermatozoide.
- La reducción del citoplasma. Se elimina el citoplasma innecesario formando un cuerpo residual, que es fagocitado por las células de Sertoli.
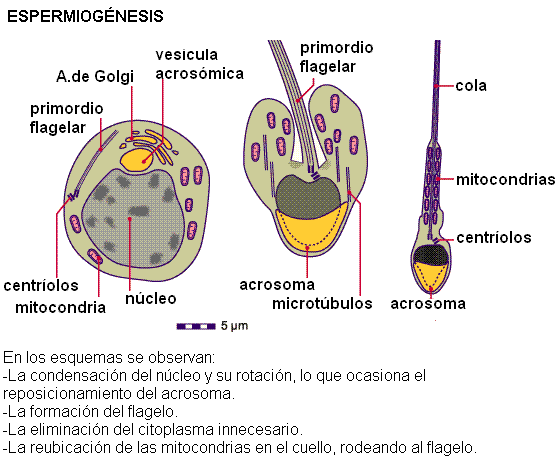 |
Una vez maduro, el espermatozoide es una célula pequeña, móvil e hidrodinámica, al servicio de la propagación del material genético paterno. Posee una cabeza ocupada casi totalmente por el núcleo, con su material genético sumamente condensado y despojada de otros organoides, un cuello donde se sitúan las mitocondrias que proveerán de energía al flagelo y una cola flagelar que le otorga movilidad.
Conductos genitales y glándulas anexas: recorrido de los espermatozoides y formación del semen Regresar
El sistema de conductos genitales permite la maduración de los espermatozoides, los almacena y los propulsa al exterior, al mismo tiempo que las glándulas anexas añaden las secreciones que componen el semen, el líquido que baña a los espermatozoides.
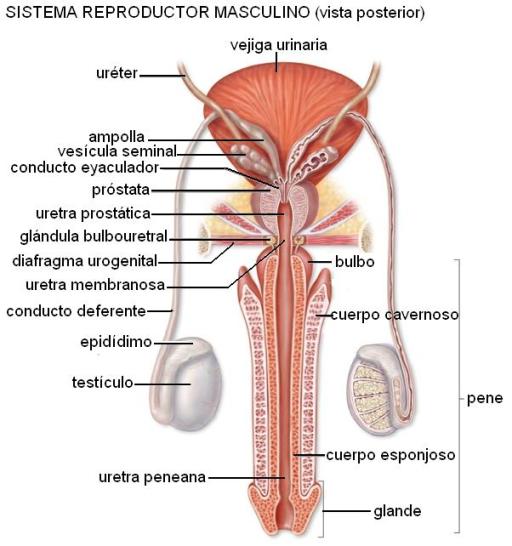
El epidídimo es un largo conducto, de unos 6m de longitud y muy enrollado, ubicado por encima del testículo. Los espermatozoides permanecen unos 12 días dentro del epidídimo, donde completan su maduración y adquieren la capacidad de moverse, haciéndose potencialmente fértiles.
El epidídimo se continúa con el conducto deferente, un tubo muscular de unos 40 cm de longitud, el cual atraviesa el conducto inguinal formando parte del cordón espermático junto con los vasos y nervios del testículo. Ya en la cavidad abdominal, el conducto deferente se une con la vesícula seminal de su lado, formando el conducto eyaculador. El conducto deferente es el principal sitio de almacenamiento de los espermatozoides.
Las vesículas seminales son pequeñas glándulas que elaboran el 60% del líquido seminal. Secretan un líquido mucoide, que contiene fibrinógeno y es rico en fructosa, destinada a nutrir a los espermatozoides. En los fluidos provenientes de las vesículas seminales también hay abundantes prostaglandinas. Se cree que éstas serían responsables de inducir una peristalsis invertida en el tracto genital femenino, favoreciendo así el ascenso de los espermatozoides por el mismo.
Los conductos eyaculadores derecho e izquierdo conducen el semen hasta la uretra, donde desembocan en el espesor de la próstata.
La próstata es una pequeña glándula que rodea a la uretra cuando ésta abandona la vejiga urinaria. Aporta alrededor de un 20-30% del semen. Secreta un líquido lechoso y alcalino, que contrarresta la acidez de las secreciones producidas por el conducto deferente y las vesículas seminales, lo mismo que la acidez vaginal. La secreción prostática contiene fosfato, bicarbonato, citrato, una enzima de la coagulación y profibrinolisina.
La próstata tiende a hipertrofiarse en los adultos, pudiendo obstruir la vía urinaria. También es un lugar frecuente de carcinogénesis en el varón.
La parte final de la vía espermática la constituye la uretra, órgano común a los sistemas urinario y genital masculinos. La uretra presenta tres porciones: prostática, membranosa y peniana. La uretra membranosa recibe las secreciones mucosas de las glándulas de Cowper o bulbouretrales. En la uretra peniana también se vuelca el moco secretado por las glándulas uretrales. La uretra peniana termina en el meato urinario, en el extremo del pene, por donde el semen alcanza el exterior.
Los espermatozoides pueden vivir varias semanas dentro de los conductos genitales masculinos. Si no son eyaculados, se reabsorben. Una vez eyaculados sólo sobreviven entre 24 y 48 horas. En el tracto genital femenino se lleva a cabo la última etapa de la diferenciación de los espermatozoides, la capacitación, en la cual se ajustan ciertos detalles de su estructura que los habilitan para la fecundación.
Pene Regresar
El pene es el órgano encargado de transferir el semen al aparato genital femenino. Presenta una porción perineal y una porción libre y se encuentra situado por encima de las bolsas, delante de la sínfisis pubiana.
El pene está formado por los tejidos eréctiles y las cubiertas que los rodean: una envoltura fibrosa, llamada fascia peniana, músculo liso y piel.
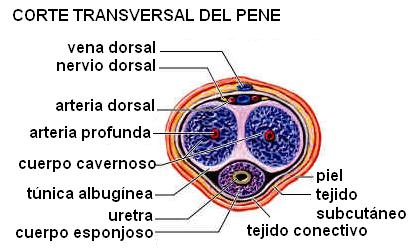 |
Los tejidos eréctiles incluyen los cuerpos cavernosos, el cuerpo esponjoso, dentro del cual transcurre la uretra, y el glande, que forma el extremo anterior del pene. El glande está rodeado por un repliegue tegumentario denominado prepucio.
Los tejidos eréctiles presentan espacios sinusoidales que se dilatan durante la excitación sexual y se ingurgitan con sangre. La fascia del pene se opone a la presión ejercida por la sangre y ocluye el drenaje venoso, en la zona dorsal, manteniendo la congestión sanguínea. De esta manera se producen la erección y la rigidez necesarias para que pueda tener lugar el coito.
Cuando se inicia la excitación, las glándulas bulbouretrales y uretrales secretan moco, el cual favorece la lubricación en el acto sexual.
Tanto la erección como la lubricación están controladas por el sistema nervioso parasimpático.
Emisión y eyaculación Regresar
La emisión y la eyaculación son la culminación del acto sexual masculino. La emisión es un reflejo simpático que se desencadena cuando el estímulo sexual es muy intenso. Consiste en el avance del semen hacia la uretra, provocado por la contracción secuencial de las paredes musculares del conducto deferente, la próstata y las vesículas seminales. Todas las secreciones se mezclan en la uretra con el moco previamente secretado por las glándulas bulbouretrales.
La repleción de la uretra genera señales sensitivas que ponen en marcha otro reflejo simpático. En respuesta, se producen las contracciones rítmicas de los conductos genitales y de los músculos isquiocavernoso y bulbocavernoso (en la base del pene), las que ocasionan la eyaculación o expulsión del semen a través del meato urinario.
Al período completo de emisión y eyaculación se lo denomina orgasmo masculino.
Al orgasmo sigue una fase de resolución, en la cual se revierten los cambios genitales y los cambios sistémicos (por ejemplo el aumento de la presión arterial) que acompañan a la fase de excitación.
Regulación hormonal del sistema reproductor masculino Regresar
Las hormonas sexuales masculinas, de las cuales la testosterona es la más importante, son esteroides secretados por las células de Leydig del intersticio testicular.
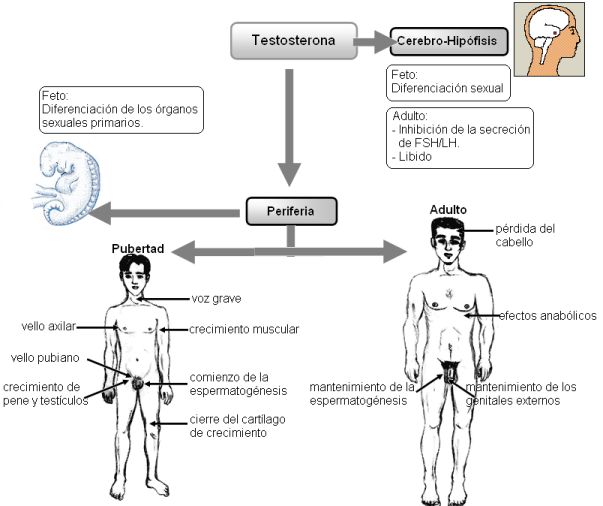
La testosterona es la responsable de la aparición de los caracteres sexuales primarios (diferenciación sexual) en los fetos portadores del cromosoma Y.
A partir de la pubertad, luego de una fase temprana de crecimiento gobernada por los esteroides de la corteza adrenal y la hormona del crecimiento, sobreviene la etapa de crecimiento acelerado (el “estirón”). Simultáneamente, la testosterona induce la aparición de los caracteres sexuales secundarios y el inicio de la fertilidad. Cuando se alcanzan los altos niveles de testosterona del adulto se produce el cierre de los cartílagos epifisarios y el cese del crecimiento.
En el adulto, la testosterona sostiene los caracteres sexuales secundarios y la espermatogénesis.
La secreción de testosterona y la función reproductiva están bajo el control del eje hipotálamo-hipofisario.
 |
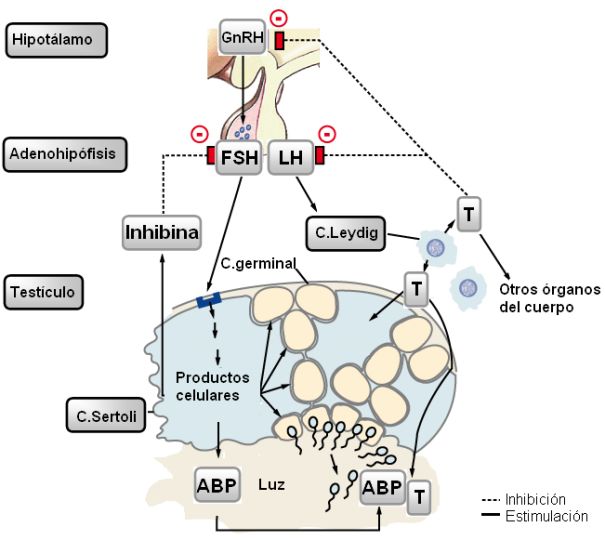
En la pubertad se produce un aumento de la secreción de factor liberador de gonadotrofinas (GnRH) por el hipotálamo. Éste estimula a la hipófisis anterior, que incrementa la secreción de las hormonas gonadotróficas: hormona folículo estimulante (FSH) y hormona luteinizante (LH).
La FSH actúa sobre las células de Sertoli del testículo, las cuales, por intermedio de sus productos celulares, promueven la espermatogénesis en los túbulos seminíferos.
Las células de Sertoli, bajo el estímulo de la FSH, también secretan proteína fijadora de andrógenos (ABP), una globulina que se une a la testosterona, manteniendo elevados niveles de la misma en la luz tubular, para beneficio de las células de la línea germinal.
Las células blanco de la LH son las células de Leydig. La LH induce la síntesis y secreción de testosterona (T) por las células de Leydig. Además de ser un factor necesario para la espermatogénesis, la testosterona ejerce efectos en muchos otros órganos, como ya se mencionó.
El equilibrio de todas las hormonas que interaccionan en la regulación de las funciones reproductivas masculinas es controlado por varios mecanismos de retroalimentación negativa.
En primer lugar, hay un mecanismo corto, en el cual elevados niveles de las hormonas gonadotróficas inhiben la secreción del factor liberador de gonadotrofinas en el hipotálamo.
Por otra parte, existe una retroalimentación entre las hormonas testiculares y el eje hipotálamo- hipófisis. La testosterona inhibe la secreción de LH. La inhibina, hormona producida por las células de Sertoli, inhibe la secreción de FSH.
 |
Sistema reproductor femenino Regresar
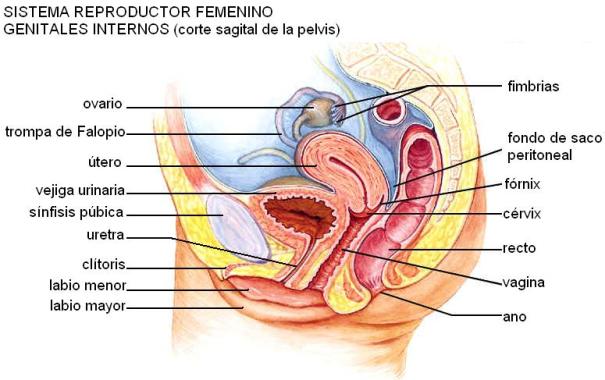
El sistema reproductor femenino está formado por los órganos reproductores o genitales externos e internos. Los primeros reciben, en conjunto, el nombre de vulva. Los genitales internos son los ovarios, las trompas de Falopio, el útero y la vagina.
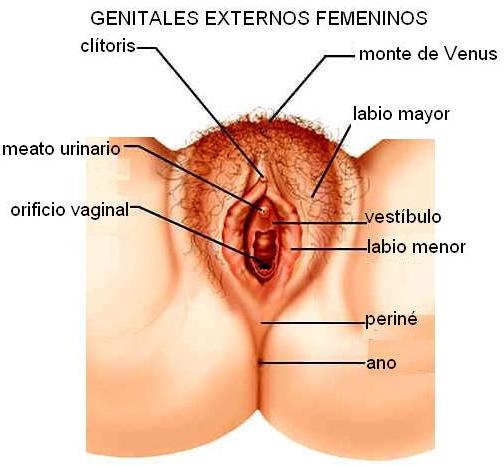
Genitales externos femeninos Regresar
La vulva está formada por varias estructuras que rodean la entrada de la vagina. Comprende los labios mayores, los labios menores y el clítoris.
Los labios mayores son dos repliegues cutáneos que ocupan la parte externa de la vulva. Por delante forman una almohadilla de grasa llamada monte de Venus, situada sobre el pubis. Por detrás, entre la vagina y el ano, los labios mayores se unen a una serie de músculos para formar el periné, o piso de la pelvis.
Los labios menores son dos finos repliegues de piel situados por dentro de los labios mayores. Delimitan el vestíbulo, espacio donde se ubican, de adelante atrás, el meato urinario y el orificio vaginal. Este último, en la mujer virgen, está parcialmente cerrado por una membrana llamada himen. Por delante, los labios menores forman un capuchón que cubre parte del clítoris. Éste es un órgano eréctil, situado en la parte anterior y superior de la vulva.
Genitales internos femeninos Regresar
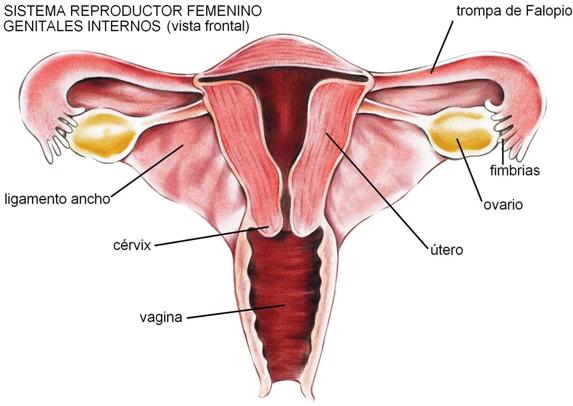 |
Los ovarios son las gónadas femeninas. Son órganos pares, del tamaño de una almendra, ubicados en la cavidad pélvica. Poseen dos funciones: la ovogénesis y la secreción de hormonas femeninas.
Las trompas de Falopio son tubos musculares que se extienden desde la extremidad externa del ovario hasta el ángulo superior del útero. Su porción externa, el pabellón, tiene la forma de un embudo y un borde irregular, formando lengüetas, que reciben el nombre de fimbrias. Internamente, las trompas están tapizadas por un epitelio ciliado y mucoso.
Las trompas transportan al ovocito liberado por el ovario. Los espermatozoides ascienden por las trompas y allí se produce la fecundación. Cuando hay fecundación, las secreciones mucosas de la trompa sirven para nutrir al embrión, mientras éste la recorre en dirección al útero.
El útero o matriz es el órgano donde se implanta y se desarrolla el embrión hasta el nacimiento.
Es un órgano hueco, muscular, en forma de pera, de tamaño ligeramente inferior al puño y está tapizado por el endometrio. Éste tiene dos capas principales: la funcional y la basal. La funcional es expulsada durante la menstruación, mientras que la basal permanece y regenera la capa eliminada. Los músculos lisos de las paredes del útero reciben el nombre de miometrio. Las contracciones del miometrio permiten la expulsión del feto y de la placenta en el momento del parto.
El extremo inferior del útero posee un esfínter muscular, el cérvix o cuello uterino, por donde pasan los espermatozoides en su camino hacia el ovocito. En el momento del nacimiento, el cuello se dilata y permite la salida del feto.
La vagina es un tubo muscular que comunica el cuello del útero con el exterior del cuerpo. Es el órgano receptivo para el pene y también el canal de parto, y su interior es ligeramente ácido.
Ciclo menstrual Regresar
| Durante la vida reproductiva de la mujer se producen variaciones cíclicas mensuales de la secreción de las hormonas que controlan al aparato reproductor, con las consecuentes modificaciones en el ovario, el útero y otros órganos sexuales, las cuales preparan al organismo para un posible embarazo. Ese patrón rítmico de cambios recibe el nombre de ciclo sexual femenino o ciclo menstrual. |
En la pubertad se experimenta la primera pérdida de sangre menstrual, o menarca, que constituye el primer signo de las modificaciones uterinas. La menopausia, alrededor de los 50 años, es el cese de los ciclos menstruales, lo que pone fin a la vida reproductiva.
Normalmente, los ciclos menstruales solo se interrumpen durante el embarazo.
La duración de los ciclos menstruales es sumamente variable; pueden ser considerados normales ciclos tan cortos, de 20 días, como ciclos largos, de hasta 45 días. La variación en la duración del ciclo menstrual es mayor en mujeres muy jóvenes y también en las de edad próxima a la menopausia.
Habitualmente se describe un ciclo tipo de 28 días. El primer día de sangrado menstrual corresponde al día 1 del ciclo, mientras que en la mitad del mismo, alrededor del día 14, tiene lugar la liberación de un ovocito secundario desde el ovario hacia la trompa de Falopio, fenómeno denominado “ovulación”.
Ciclo ovárico Regresar
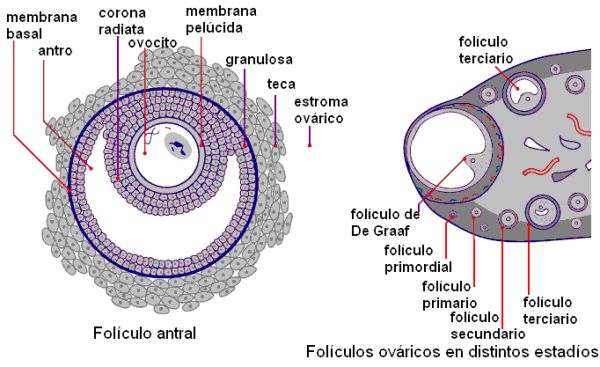
Los ovarios están formados por una zona cortical y una zona medular. En la zona cortical se ubican los folículos ováricos. En el momento del nacimiento, los folículos constan de un ovocito primario, detenido en la profase de la primera división meiótica, rodeado de una capa de células foliculares planas. Muchos de estos folículos primordiales se atrofian antes de la pubertad. Para entonces, perduran alrededor de 400.000 folículos primordiales, los cuales se han mantenido quiescentes hasta ese momento. A partir de la pubertad, bajo control hormonal, un cierto número de folículos comienza a madurar durante cada ciclo menstrual, por lo cual en el ovario maduro coexisten folículos primordiales con otros en distintas etapas de crecimiento y maduración.
Las etapas en el desarrollo folicular son las siguientes:
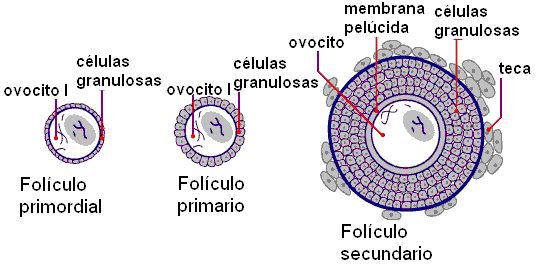
• Folículo primordial: es el folículo en la fase de reposo. El ovocito primario está rodeado por las células foliculares que reciben el nombre de “células de la granulosa”, por fuera de las cuales se encuentra una membrana basal.
• Folículo primario: se caracteriza por un aumento de tamaño del ovocito, en cuyo derredor se forma una membrana, la zona pelúcida. Las células de la granulosa se multiplican formando varias capas, y adquieren forma cúbica.
• Folículo secundario: continúa la proliferación de las células granulosas. Las células del estroma (tejido conectivo) del ovario forman la teca, alrededor de la membrana basal. Entre las células de la granulosa comienza a acumularse líquido.
 |
• Folículo terciario o antral: se forma el antro, cavidad llena de líquido, entre las células de la granulosa. La teca se diferencia en dos capas, la teca interna, con predominio celular y la teca externa, de tejido conectivo más denso.
• Folículo de De Graaf: es el folículo preovulatorio. Alcanza alrededor de 2,5 cm de diámetro y sobresale en la superficie de la corteza ovárica. El ovocito ocupa dentro del mismo una posición excéntrica. Las células de la granulosa forman un conjunto que rodea al ovocito (la corona radiata) y un grupo que lo conecta con la periferia del folículo (cúmulo oóforo).
El desarrollo de un folículo primordial hasta la fase antral dura aproximadamente 3 meses.
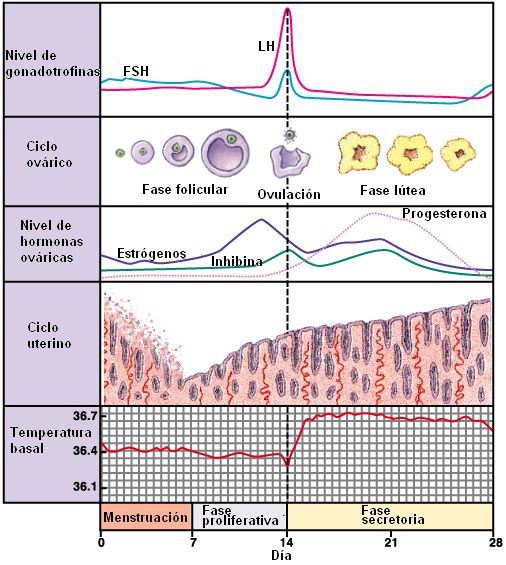
| En el ciclo ovárico se describen dos fases: la fase folicular, que se extiende desde el día 1 hasta el día 14, cuando ocurre la ovulación, y la fase lútea, que abarca desde la ovulación hasta el fin del ciclo. |
La fase folicular se inicia con el crecimiento rápido, debido al estímulo hormonal, de varios folículos. Aproximadamente el día 7, se selecciona uno de ellos, el folículo dominante, que completará su desarrollo hasta la ovulación. El resto de los folículos degenera, convirtiéndose en folículos atrésicos.
Durante el desarrollo del folículo dominante se completa la primera división meiótica, dando lugar a la formación del ovocito II y el primer corpúsculo polar, que permanece en el espacio perivitelino (entre el ovocito y la membrana pelúcida).
El día 14, el folículo dominante se rompe en la superficie del ovario y se concreta la ovulación. El ovocito II es liberado del ovario junto con la corona radiata y el líquido folicular, dirigiéndose hacia la trompa de Falopio, donde inicia la segunda división meiótica.
Si en las 24 horas precedentes o en las 24 horas siguientes a la ovulación se produjo una relación sexual, de manera que en el tracto genital femenino se encuentren espermatozoides viables, entonces el ovocito II podrá ser fecundado y completará su segunda división meiótica, con la inmediata formación de la cigota. Si no se produce la fecundación, el ovocito II muere en la trompa de Falopio.
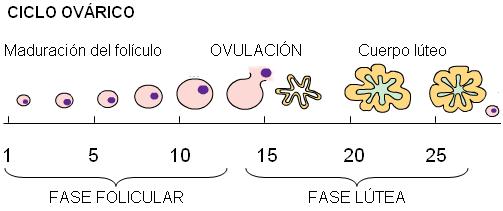 |
Después de la ovulación, los restos del folículo que permanecen en el ovario se transforman en el cuerpo amarillo o cuerpo lúteo. Las células de la granulosa y de la teca interna se diferencian, adquieren un gran desarrollo del retículo endoplasmático liso, acumulan lípidos (lo que les da el color amarillo o lúteo) y se especializan en la secreción de hormonas sexuales femeninas, de naturaleza esteroide. El cuerpo lúteo tiene una vida previamente programada, de aproximadamente 10 días. Si no ocurre la fecundación, después de ese lapso el cuerpo lúteo involuciona y se convierte en el cuerpo blanco, inactivo, que puede persistir unos meses hasta desaparecer por completo.
Cuando se produce la fecundación, sin embargo, el cuerpo lúteo perdura hasta el 3º o 4º mes de embarazo.
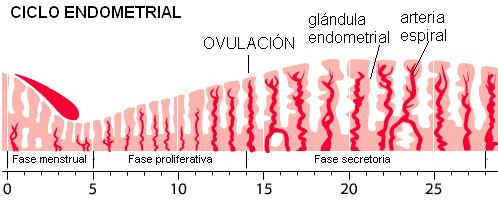
Ciclo endometrial Regresar
El útero es el órgano donde se implanta y desarrolla el embrión cuando se produce un embarazo. Los cambios que afectan al endometrio durante un ciclo menstrual lo preparan para recibir al embrión después de la fase ovulatoria. Cuando no hay fecundación, el cuerpo amarillo involuciona y la falta de estímulo hormonal provoca la caída de la capa funcional del endometrio.
Las fases del ciclo endometrial son:
• Fase menstrual: las arterias espirales, que nutren al estrato funcional del endometrio, sufren una vasoconstricción, alrededor del día 25 ó 26 del ciclo. La falta de irrigación ocasiona la isquemia (carencia de oxígeno) y necrosis (muerte) del endometrio funcional. Finalmente, éste se descama y se elimina junto con sangre en la que se conoce como fase menstrual o menstruación. El primer día de eliminación del flujo menstrual se considera el día 1 del ciclo. La menstruación se extiende por 5 días, aproximadamente.
• Fase proliferativa: durante esta etapa se regenera el estrato funcional del endometrio a partir de la capa basal. Las glándulas endometriales proliferan. Para el día 14, cuando tiene lugar la ovulación, el endometrio está completamente regenerado.
• Fase secretoria: corresponde a un mayor desarrollo y actividad de las glándulas endometriales. Las células glandulares se llenan de gránulos de glucógeno. El pico de actividad glandular se alcanza hacia el día 21. Éste es el tiempo en que se inicia la implantación del embrión, en caso de haber ocurrido la fecundación.
Otros cambios en el ciclo menstrual Regresar
Además de las modificaciones a nivel del ovario y el endometrio, otros cambios se repiten cíclicamente en el organismo femenino. Por ejemplo, varía la distribución de los diversos tipos celulares en el epitelio vaginal. El cuello del útero produce un moco acuoso, ideal para la penetración del esperma, en la fase folicular tardía, cuando se acerca la ovulación. En cambio, durante la fase lútea, el moco cervical es más escaso y viscoso.
Regulación hormonal del sistema reproductor femenino Regresar
El sistema reproductor femenino y el ciclo menstrual se hallan, al igual que el sistema reproductor masculino, bajo el control de las hormonas del eje hipotálamo-hipofisario. Existen tres jerarquías de hormonas:
1- Hormona liberadora de gonatrofinas (GnRH) hipotalámica. Es la encargada de estimular la secreción de gonadotrofinas por la hipófisis. Sus niveles aumentan en la pubertad y disminuyen en la menopausia. La secreción de GnRH está sujeta a retroalimentación negativa por los esteroides ováricos.
2- Gonadotrofinas hipofisarias: FSH y LH. La FSH induce la maduración del folículo ovárico. La LH dispara la ovulación e induce la secreción de hormonas sexuales por el ovario. Niveles moderados de esteroides ejercen una retroalimentación negativa de la secreción de gonadotrofinas. En cambio, altos niveles de estrógenos estimulan la secreción de gonadotrofinas hipofisarias. La liberación de FSH es inhibida por la inhibina, proveniente del ovario.
3- Hormonas sexuales femeninas: estrógenos y progestágenos. Ambos provienen del ovario y son de naturaleza esteroide. Los estrógenos (principalmente estradiol) determinan la aparición de los caracteres sexuales secundarios en la pubertad y estimulan la proliferación del endometrio en cada ciclo menstrual. La progesterona (el progestágeno más potente) prepara al endometrio para la implantación.
Regulación hormonal del ciclo menstrual Regresar
Durante la niñez, los folículos primordiales se mantienen en reposo. Se cree que esto es debido a un factor inhibidor de la maduración del ovocito, secretado por las células de la granulosa.
Después, tras la pubertad, la FSH y la LH se secretan en grandes cantidades, estimulando el crecimiento del ovario y la maduración de los folículos.
A poco de iniciado cada ciclo menstrual, las concentraciones de gonadotrofinas van en aumento, la de FSH precediendo a la de LH. El aumento en los niveles de FSH causa un crecimiento acelerado de entre 6 y 12 folículos, los que se desarrollan en folículos antrales.
Una vez formados los folículos antrales, el crecimiento de éstos es aún más acelerado. Este crecimiento es causado por la secreción de estrógenos, secretados por las células de la granulosa y de la teca interna, bajo el estímulo de la FSH. Los estrógenos se acumulan en el folículo, formando el líquido que ocupa el antro, e inducen un aumento en el número de receptores de FSH. Así se produce una retroalimentación positiva, ya que ahora los folículos antrales se hacen más sensibles a los efectos de la FSH. Además, la acción combinada de la FSH y los estrógenos promueve la aparición de receptores de LH en las células de la granulosa y la teca interna.
Uno de los folículos responde más que el resto a la FSH y, debido a la retroalimentación positiva causada por los estrógenos, se convierte en el folículo dominante. Éste comienza a producir inhibina, hormona que, junto con el creciente nivel de estrógenos, inhibe la secreción de FSH. La disminución en la secreción de FSH bloquea el desarrollo de otros folículos menos adelantados, que aún no han empezado su retroalimentación positiva. Estos folículos se vuelven atrésicos, en tanto el dominante es el único que continúa su maduración.
En el folículo dominante se completa la primera división meiótica. No obstante, para el crecimiento final del folículo y la ovulación, se requiere un aumento de la LH. Un pico de LH y de FSH se produce alrededor del día 12 y ambas hormonas actúan sinérgicamente provocando la hinchazón y ruptura del folículo, con la consiguiente liberación del ovocito secundario.
El pico de LH ocasiona una disminución en la secreción de estrógenos y la luteinización (conversión a cuerpo lúteo) en los restos del folículo. El cuerpo lúteo secreta grandes cantidades de hormonas sexuales, principalmente progesterona, pero también estrógenos e inhibina.
Las hormonas ováricas inhiben la secreción de FSH y LH, cuyos niveles decaen en la segunda mitad del ciclo. Como el cuerpo lúteo depende del estímulo de la LH, comienza a involucionar 6 a 8 días después de la ovulación, a menos que haya ocurrido la fecundación.
Con la involución del cuerpo lúteo también decaen los niveles de progesterona y estrógenos. Esto provoca dos efectos. A nivel del endometrio, causa la necrosis, pues aquél se sostiene gracias a las hormonas ováricas. A nivel del eje hipotálamo hipofisario, merma la inhibición de la secreción de gonadotrofinas. Como resultado, el endometrio cae, y se inicia un nuevo ciclo.
Cuando hay fecundación, el embrión secreta gonadotrofina coriónica humana, hormona que sostiene al cuerpo lúteo. De esta forma el endometrio se mantiene, permitiendo la implantación del embrión.
 |
Efectos de los esteroides sexuales: estrógenos y progestágenos Regresar
Estrógenos |
| En la pubertad |
Desarrollo del útero y las mamas.
Depósito de grasa que caracteriza al cuerpo femenino (en glúteos y muslos).
Aumento de la actividad osteoblástica y cierre del cartílago de crecimiento. |
| En cada ciclo menstrual |
Proliferación del endometrio.
Aclaramiento del moco cervical. |
| En el embarazo |
Aumento del flujo sanguíneo uterino.
Hipertrofia del miometrio.
Crecimiento de los conductos mamarios.
Retención de líquido. |
| En la menopausia |
La disminución en la producción de estrógenos ocasiona:
Cese de la ovulación.
Sequedad de la piel.
Osteoporosis.
Atrofia de las mamas y la vagina. |
Progestágenos |
| SNC |
Termogénesis.
Inhibición de la secreción de GnRH.
Estimulación de la libido. |
| Mamas |
Limita la lactancia preparto.
Lactogénesis. |
| Trompa de Falopio |
Estimula el transporte de gametas. |
| Útero |
Crecimiento y desarrollo del endometrio.
Acumulación de glucógeno en el endometrio.
Secreción en las glándulas uterinas.
Disminuye la sensibilidad del miometrio a la oxitocina. |
Fecundación Regresar
La fecundación o fertilización es la fusión de los gametos femenino y masculino, lo que da por resultado la formación de la célula huevo o cigota.
Cuando el semen es eyaculado dentro de la vagina, forma un débil coágulo en las partes profundas de la misma, donde se encuentra el cérvix. En los primeros minutos tras la eyaculación, los espermatozoides permanecen relativamente inmóviles. Luego el coágulo se disuelve, gracias a la fibrinolisina prostática. A medida que se disuelve el coágulo, los espermatozoides adquieren mayor movilidad.
A pesar de su maduración en el epidídimo, factores inhibidores presentes en los conductos masculinos hacen que los espermatozoides no estén completamente aptos para fecundar, sino hasta que toman contacto con los líquidos del sistema reproductor femenino. Allí se produce la capacitación, una serie de cambios que activan a los espermatozoides, haciendo posible la fecundación.
En el tracto genital femenino, los espermatozoides pierden colesterol de sus membranas y se hacen más permeables al calcio. El ingreso de calcio ocasiona la aparición de movimientos enérgicos del flagelo (hiperactivación).
El encuentro con el ovocito II, detenido en la metafase de la segunda división meiótica, se produce en el tercio distal de la trompa de Falopio.
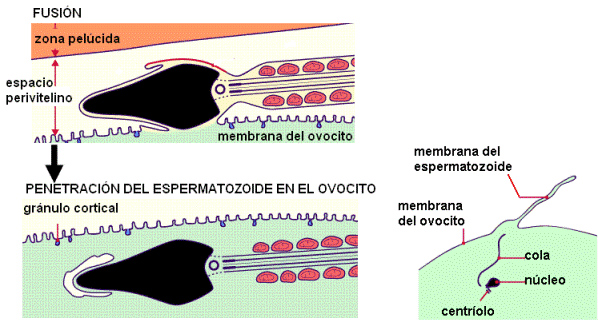
Los espermatozoides avanzan a través de la corona radiata, abriéndose paso mediante la hialuronidasa que poseen en sus membranas; ésta digiere el material cementante que une a las células foliculares.
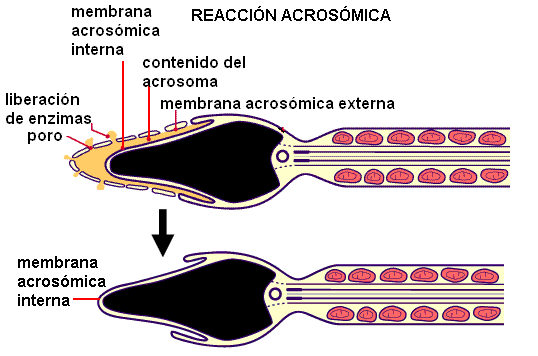
Cuando el espermatozoide alcanza la membrana pelúcida se produce la reacción acrosómica. Dicha reacción consiste en la fusión de la membrana acrosómica externa con la membrana plasmática del espermatozoide, y es disparada por el contacto con una proteína de la membrana pelúcida. La reacción acrosómica libera más enzimas que separan la corona radiata, y deja expuesta la membrana interna del acrosoma, que cubre la cabeza del espermatozoide.
 |
 |
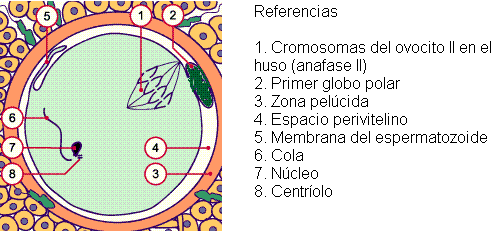
Posteriormente, ocurre la fusión entre la membrana del espermatozoide y la membrana del ovocito. La fusión de las membranas permite el ingreso de lo que resta de la cabeza, el cuello y la cola del espermatozoide, al citoplasma del ovocito. Sin embargo, solamente persisten el ADN y el centríolo, ya que las otras partes desaparecen rápidamente.
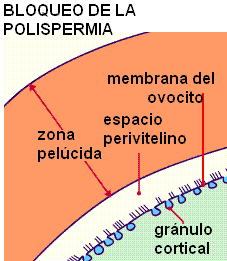
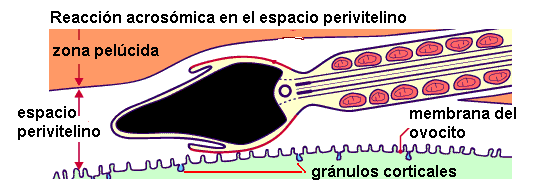
Después de la fusión con un espermatozoide, el ovocito experimenta cambios que bloquean la polispermia, es decir, el ingreso de otros espermatozoides. Gránulos corticales, ubicados por debajo de la membrana plasmática del ovocito, se exocitan: es la llamada reacción cortical. Las enzimas exocitadas inmovilizan a los espermatozoides que quedan atrapados en la zona pelúcida. Además, la unión de los gránulos a la membrana plasmática añade, a esta última, proteínas que impiden una nueva fusión.
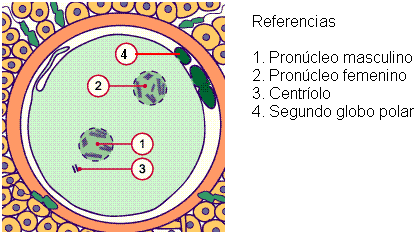
Simultáneamente con el bloqueo de la polispermia, el ovocito II reanuda la segunda división meiótica, formando dos núcleos haploides: el del óvulo y el del segundo cuerpo polar.
El núcleo del óvulo y el del espermatozoide, llamados pronúcleos femenino y masculino, descondensan y duplican su material genético.
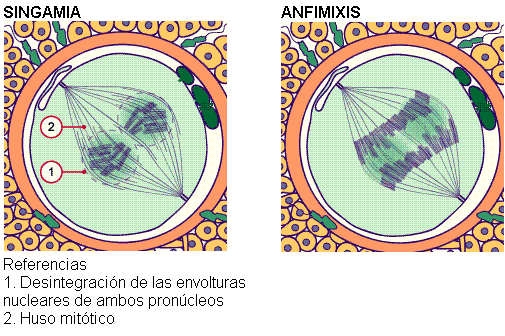
A continuación, las respectivas envolturas nucleares se desintegran, produciéndose la singamia o fusión de ambos pronúcleos. Así surge la cigota.
Inmediatamente, los cromosomas maternos y paternos de la cigota se alinean en el plano ecuatorial (anfimixis), iniciando la primera de innumerables divisiones mitóticas que darán origen a un nuevo organismo.
|
|