Capítulo 5: Medio Interno
Los sistemas de órganos Regresar
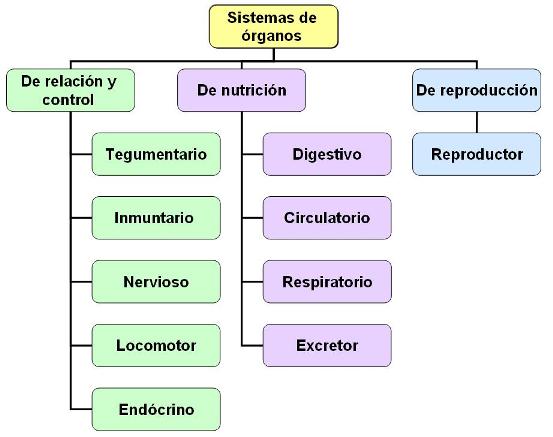
Los organismos animales están organizados en aparatos o sistemas de órganos, encargados de realizar las grandes funciones que un ser vivo requiere para mantenerse y reproducirse.
Las funciones de relación y control son las que le permiten al organismo interactuar con el medio, captar estímulos, responder a ellos y ajustar las respuestas a las condiciones externas e internas. Al mismo tiempo, debe existir una coordinación de las funciones de los distintos sistemas, para mantener un equilibrio interno u homeostasis. Los sistemas de relación y control son:
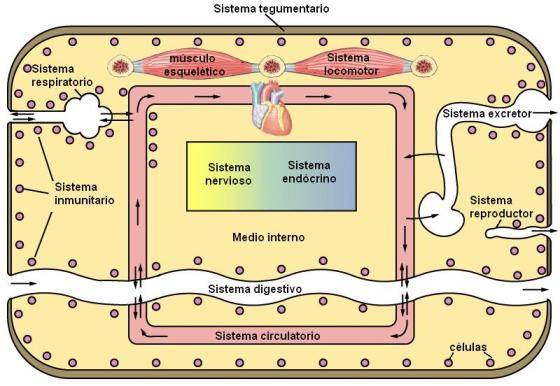
- El sistema tegumentario: es el límite del cuerpo y el que lo conecta directamente con el medio.
- El sistema inmunitario: lleva a cabo las respuestas defensivas del organismo frente a los microorganismos patógenos.
- El sistema locomotor: encargado de los movimientos.
- El sistema nervioso: coordina los estímulos con las respuestas, tanto en la relación con el medio externo como entre los distintos sistemas corporales.
- El sistema endócrino: regula funciones metabólicas, de crecimiento y reproductivas.
Las funciones de nutrición abarcan el conjunto de medios por los cuales el organismo se aprovisiona de materia y energía para la asimilación y la realización de trabajos celulares. Las funciones de nutrición involucran a los siguientes sistemas:
- Sistema digestivo: incorpora los nutrientes.
- Sistema circulatorio: transporta los nutrientes y los desechos metabólicos.
- Sistema respiratorio: intercambia gases con el medio. Incorpora oxígeno, necesario para la oxidación de los nutrientes y elimina dióxido de carbono, desecho producido durante la oxidación.
- Sistema excretor: elimina ciertos desechos del metabolismo.
Por último, el sistema reproductor está especializado en la formación de gametas y las demás funciones que posibilitan la perpetuación de la especie.
Definición de medio interno Regresar
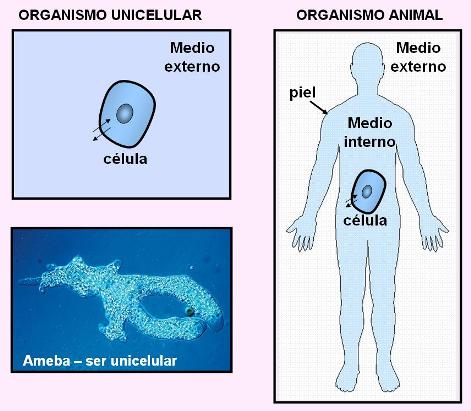
Los seres vivos aparecieron y evolucionaron en el mar. El medio acuoso que rodea a las células forma parte funcional de ellas. Para un organismo unicelular, el agua se distribuye en dos medios: uno intracelular y otro exterior. En cambio, las células que constituyen el cuerpo de un animal viven en un “mar interior”, que son los líquidos corporales. Al conjunto de los líquidos corporales que rodean a las células y están encerrados dentro de la piel de un animal se les dio el nombre de “medio interno”.
En el medio interno o líquido extracelular (LEC) de un animal se encuentran los iones y nutrientes que necesitan las células para mantener su vida. Las células serán capaces de vivir, crecer y desarrollar sus funciones especiales en tanto dispongan de las concentraciones correctas de oxígeno, glucosa, iones, aminoácidos y otros constituyentes en el medio interno. Los sistemas de nutrición, como el digestivo, el respiratorio y el excretor, son los que median los intercambios entre el medio externo y el medio interno.
 |
Compartimientos de los líquidos corporales Regresar
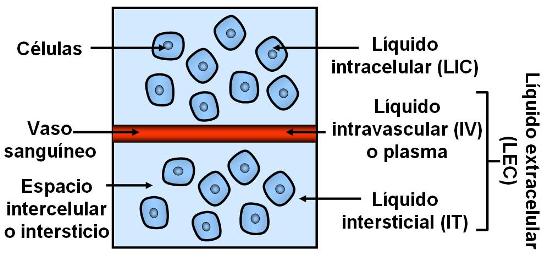
La mayor parte del agua corporal total (ACT), aproximadamente dos tercios, se encuentra en el compartimiento intracelular (LIC = líquido intracelular). El tercio restante corresponde al medio interno o líquido extracelular (LEC).
El LEC está dividido en dos compartimientos: el intravascular (IV) y el intersticial (IT).
El compartimiento intravascular corresponde al interior de los vasos sanguíneos; allí circula la sangre, cuya mayor parte está formada por un líquido llamado plasma. El plasma representa la cuarta parte del LEC.
El líquido intersticial (IT) es la porción del LEC que baña a las células y se encuentra fuera de los vasos. Éste representa las tres cuartas partes del LEC.
| Distribución del ACT en los distintos compartimientos |
| LIC |
2/3 |
| LEC |
1/3 |
LIT |
3/4 |
| Plasma |
1/4 |
Los líquidos de los diferentes compartimientos están separados por membranas biológicas:
- la membrana plasmática separa al LIC del IT y
- el epitelio que forma la pared de los vasos sanguíneos (endotelio) separa al líquido IT del plasma.
Los compartimientos donde se reparten los líquidos corporales presentan diferentes concentraciones de iones y moléculas. Las diferencias entre los compartimientos son mantenidas por la permeabilidad selectiva de las membranas.
Para comprender cómo se generan y mantienen estos desequilibrios en los líquidos corporales, esenciales para el funcionamiento orgánico, es necesario conocer algunos principios físico-químicos que rigen el movimiento de las partículas en solución y los mecanismos de transporte de las membranas biológicas.
Concepto de difusión Regresar
Cuando las partículas (moléculas o iones) están en solución en un medio líquido o gaseoso se mueven de acuerdo con la energía cinética que poseen. Los movimientos ocurren al azar, sin seguir ninguna dirección particular. Cada partícula tiene la misma probabilidad de moverse en cualquier dirección del espacio.
Si existe una zona del sistema donde la concentración del soluto es mayor, entonces es más probable que las partículas se muevan desde esa zona hacia otra, que a la inversa, por el simple hecho de que allí son más. Se registrará entonces un movimiento neto de soluto desde la zona de mayor a la de menor concentración. A ese movimiento se lo denomina difusión.
A la diferencia de concentración de un soluto a lo largo de una distancia se le da el nombre de gradiente de concentración. Cuando una partícula difunde (va desde donde está más hacia donde está menos concentrada) se dice que se mueve “a favor de gradiente”.
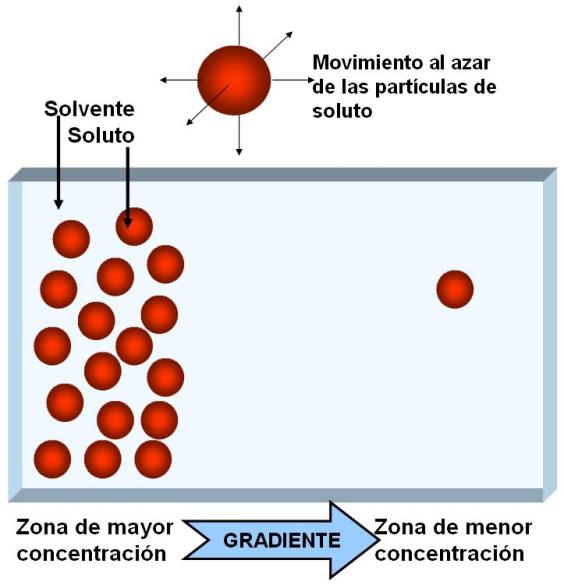 |
|
Si transcurre un tiempo, las partículas llegarán a igualar su concentración en todos los puntos del sistema; es decir, se anulará el gradiente. Las partículas seguirán con sus movimientos aleatorios después de que haya desaparecido el gradiente. Sin embargo, esos movimientos no representarán cambios en las concentraciones, pues por cada partícula que se mueva desde A hacia B habrá otra que lo haga desde B hacia A. En esa situación se habrá alcanzado un equilibrio dinámico, sin que se registre movimiento neto de partículas.
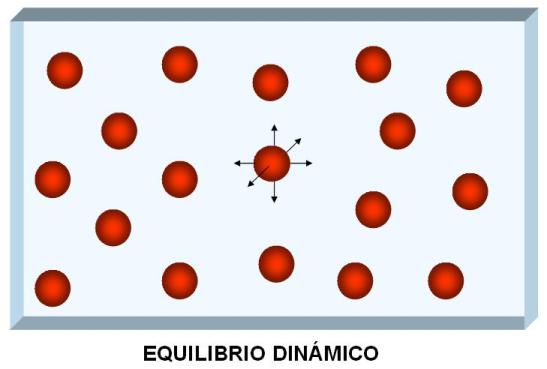 |
La difusión es un fenómeno pasivo y espontáneo que se produce en cualquier medio donde haya partículas disueltas. Cotidianamente somos testigos de procesos de difusión. Podemos observarla, por ejemplo, cuando el azúcar se reparte uniformemente en el agua de una taza, cuando el té colorea el agua en la que sumergimos el saquito o cuando al destapar un frasco con perfume, las moléculas de perfume llegan hasta nuestras fosas nasales. Todos ellos son movimientos a favor de gradiente. Los movimientos contra gradiente (desde la zona de menor a la de mayor concentración) en cambio, no son procesos espontáneos ni pasivos; son fenómenos diferentes de la difusión, para los cuales se requiere un aporte de energía.
La velocidad de difusión es directamente proporcional a la temperatura, a la solubilidad de las partículas en el medio y al gradiente de concentración, y es inversamente proporcional a la distancia a recorrer. Si los solutos son iones, la difusión ya no dependerá solamente del gradiente químico, sino también de la diferencia de carga o gradiente eléctrico. Los aniones se moverán hacia la zona más positiva y los cationes hacia la zona más negativa.
Difusión a través de la membrana plasmática Regresar
La membrana plasmática está interpuesta entre el medio intracelular y el líquido intersticial. Ambos compartimientos son soluciones acuosas con muchos tipos de iones y moléculas en solución.
Los solutos tienden a difundir espontáneamente desde un compartimiento al otro siguiendo sus respectivos gradientes electroquímicos.
Cuando un soluto difunde libremente a través de la membrana, se iguala la concentración del mismo en los medios intracelular y extracelular. La dirección del movimiento depende exclusivamente del gradiente. Por tratarse de un fenómeno espontáneo, este tipo de transporte no implica un gasto energético para la célula. Se dice por esto que la difusión es un transporte pasivo.
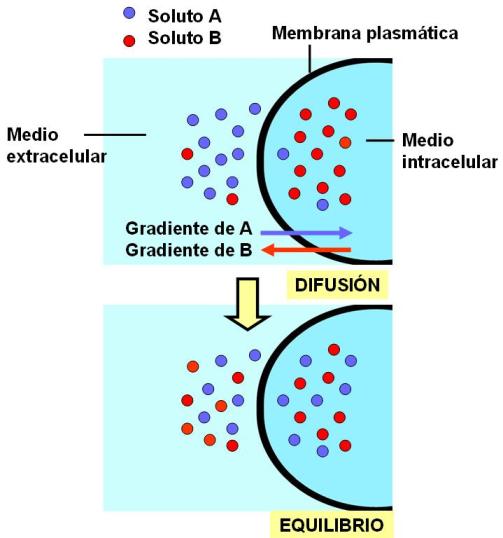 |
Cada soluto difunde independientemente de otros, a favor de su gradiente |
Sin embargo, no todos los solutos difunden libremente, pues la membrana se interpone como una barrera de permeabilidad selectiva.
La posibilidad de difusión a través de la membrana (cuando está establecido un gradiente a ambos lados de la misma) depende de dos factores:
- el tamaño de las partículas del soluto, y
- la afinidad entre el soluto y los componentes de la membrana.
En lo que respecta al tamaño, solo podrán difundir iones o moléculas relativamente pequeñas, cuyo tamaño les permita atravesar los canales que se forman entre los componentes de la membrana o por el interior de ellos.
En lo relativo a la afinidad por los componentes de membrana, los solutos se comportan de distinta forma, de acuerdo a su carácter apolar (hidrofóbico) o polar (hidrofílico). Los primeros se mueven a través de la bicapa lipídica, mientras que los segundos en general requieren la presencia de proteínas transportadoras.
Difusión simple Regresar
Es el pasaje a favor de gradiente a través de los espacios generados entre los lípidos que forman la bicapa.
La velocidad de difusión simple se ve afectada por la fluidez o libertad de movimiento que tienen los lípidos dentro de la bicapa. Cuanto más fluida es la membrana, con mayor velocidad se produce la difusión simple. La fluidez de la bicapa, a su vez, crece con el grado de insaturación de los lípidos (ver cap. 2).
Dos compuestos pequeños y apolares que se mueven por difusión simple a través de la membrana son el oxígeno (O2) y el dióxido de carbono (CO2). El oxigeno se halla dentro de las células a baja concentración, ya que es consumido continuamente en el proceso de respiración celular. Eso hace que se mantenga un gradiente con respecto al medio extracelular y el oxígeno ingresa a la célula por difusión simple. Lo contrario ocurre con el dióxido de carbono, que es generado en la respiración celular. Como su concentración en las células es mayor, el gradiente lo lleva hacia el medio extracelular.
Otros compuestos hidrofóbicos (liposolubles) de mayor tamaño, como ácidos grasos y esteroides, se mueven a través de las bicapas por difusión simple.
Algunas moléculas polares sin carga, muy pequeñas, como la urea, el etanol o el agua, pueden atravesar la bicapa lipídica, pero lo hacen en menor grado y más lentamente que las moléculas apolares.
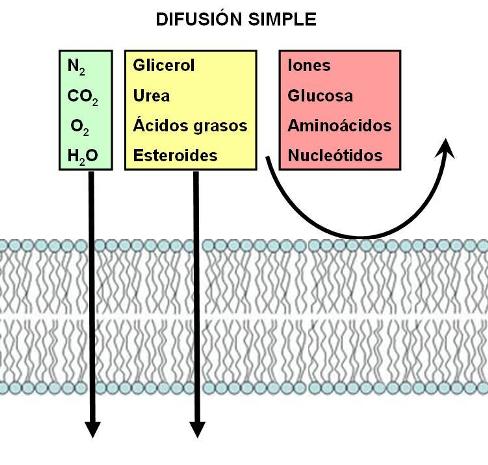 |
Difusión facilitada Regresar
Los iones y moléculas polares más grandes también pueden difundir a través de la membrana. Sin embargo, debido a su naturaleza hidrofílica, resultan rechazados por la bicapa lipídica. Este tipo de partículas ingresa a la célula o egresa de ella (según sus respectivos gradientes) a través de proteínas de la membrana llamadas proteínas transportadoras. Las proteínas transportadoras delimitan espacios rodeados por sus cadenas polares, que permiten a otras moléculas polares atravesar la membrana sin tomar contacto con el espesor hidrófobo de la bicapa.
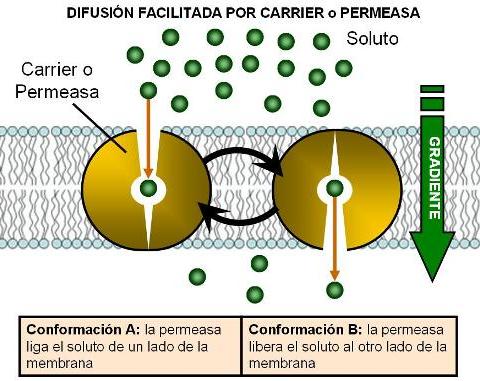
La difusión a través de proteínas transportadoras se denomina difusión facilitada.
Dos grupos de proteínas transportadoras intervienen en la difusión facilitada: los canales y los carriers o permeasas. Los canales son proteínas que ofrecen un canal hidrofílico para el pasaje de iones. Algunos canales están permanentemente abiertos, mientras que otros poseen compuertas cuyo cierre y cuya apertura están regulados por algún tipo de señal.
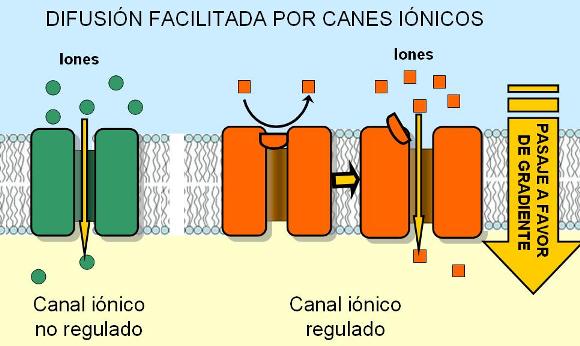 |
Los carriers o permeasas son proteínas con un sitio específico donde encaja un determinado tipo de soluto. La unión del soluto específico provoca un cambio en la conformación del carrier. Este cambio conformacional arrastra al soluto hacia el lado opuesto de la membrana, donde es liberado. Después de liberar el soluto, el carrier retoma su conformación inicial.
En la membrana plasmática existen carriers para transportar glucosa, aminoácidos, dipéptidos, y otras moléculas polares.
Ósmosis. Transporte de agua a través de la membrana plasmática Regresar
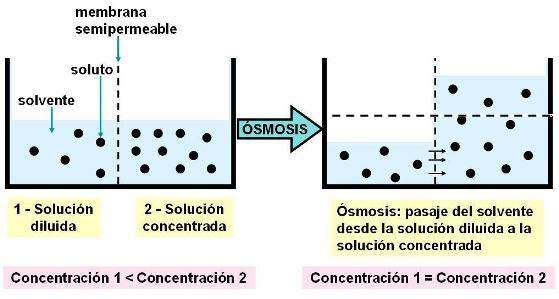
La ósmosis se define como el pasaje de un solvente, a través de una membrana semipermeable, desde la solución más diluida a la más concentrada.
Hay situaciones en las cuales la membrana que separa dos compartimientos permite el pasaje del solvente, pero no el del soluto (membrana semipermeable). Si las soluciones ubicadas a ambos lados de la membrana presentan diferentes concentraciones de soluto, éstos tenderán a difundir, pero no podrán hacerlo porque la membrana no es permeable para ellos. Como las partículas disueltas ejercen atracción sobre el solvente, es éste el que se mueve de un compartimiento al otro. El solvente se mueve hacia la solución más concentrada, debido a que es la que tiene, relativamente, más partículas de soluto y por consiguiente ejerce mayor atracción.
A la solución que es más concentrada con respecto a otra se la llama hiperosmótica; la menos concentrada es la hipoosmótica.
A medida que ocurre la ósmosis, la solución diluida se concentra y la concentrada se diluye. La ósmosis tiende a igualar la concentración en ambos medios. Cuando dos medios tienen la misma concentración son isoosmóticos uno respecto del otro. Una vez igualada la concentración, se alcanza un equilibrio dinámico: por cada molécula de solvente que pasa hacia un lado de la membrana, otra lo hará hacia el lado opuesto.
La ósmosis no iguala la cantidad de soluto y solvente en ambos medios, sino que iguala la relación entre uno y otro, es decir: la concentración. En la solución hiperosmótica se produce un aumento del volumen, mientras que en la hipoosmótica, el volumen disminuye.
 |
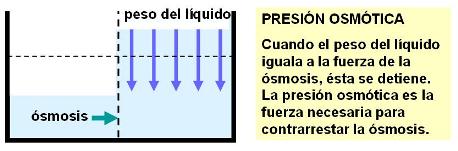
La ósmosis puede detenerse antes de alcanzarse el equilibrio de concentración, porque alguna otra fuerza se opone a ella. Por ejemplo, la ósmosis puede ser limitada porque el compartimiento hacia donde va el solvente no tiene más capacidad. O puede ser contrarrestada por el mismo peso de la columna líquida.
Se define a la presión osmótica como la presión necesaria para contrarrestar la ósmosis. La presión osmótica de una solución mide, indirectamente, cuán fuerte es la atracción que la solución ejerce sobre el solvente.
 |
La presión osmótica de una solución no depende del tipo de partículas disueltas, sino solamente de la cantidad de las mismas. Para medir la cantidad de partículas disueltas osmóticamente activas en una solución se utiliza la osmolaridad. En fisiología la osmolaridad se mide en miliosmoles (1 mOsm = 10-3 osmoles).
La membrana celular se comporta, para muchas sustancias, como una membrana semipermeable. El movimiento de agua entre el líquido extracelular y el líquido intracelular se produce por ósmosis. Por lo tanto, el ingreso de agua a la célula y su egreso siguen los movimientos de los solutos.
La ósmosis es un transporte pasivo, que no requiere aporte de energía por parte de la célula. El agua, pequeña molécula polar sin carga, como ya se mencionó, puede atravesar la bicapa lipídica.
Transporte activo Regresar
Las células son capaces de transportar solutos en contra de su gradiente de concentración (desde la zona de menor a la de mayor concentración). Esto les permite establecer un desequilibrio entre las concentraciones de algunas sustancias en el líquido intracelular y el líquido extracelular.
Los transportes contra gradiente requieren:
- la presencia de una proteína transportadora, y
- una fuente de energía.
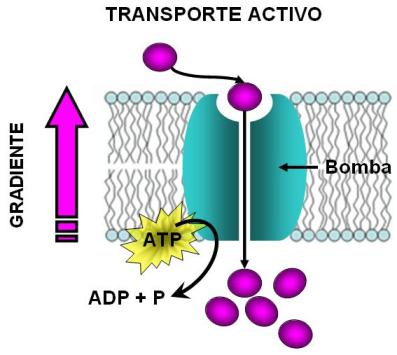
A los transportes realizados contra gradiente, por intermedio de proteínas transportadoras y con gasto de energía se les da el nombre de transporte activo.
Un tipo de transporte activo, llamado transporte activo primario, es el que está mediado por bombas. Las bombas son proteínas transportadoras específicas para determinado tipo de soluto. Pueden captar el soluto de un lado de la membrana, donde se halla a menor concentración y soltarlo del lado opuesto, donde su concentración es mayor. El pasaje del soluto a través de la bomba se debe a un cambio conformacional de la proteína. Para concretar este cambio conformacional y por ende el transporte de soluto, la bomba depende de la presencia de ATP.
La molécula de ATP es hidrolizada durante el transporte, de manera que libera la energía que tiene almacenada. Esta energía se utiliza para propulsar el trabajo que implica mover un soluto en contra de su gradiente.
 |
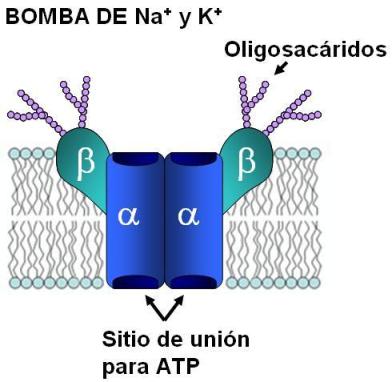
Bomba de sodio y potasio (Na+/K+ ATPasa) Regresar
La bomba de sodio (Na+) y potasio (K+) se encuentra en las membranas de las células animales. Es una proteína integrada por cuatro subunidades, dos alfa y dos beta (alfa2-beta2). Las subunidades beta poseen varias cadenas oligosacarídicas hacia la cara extracelular. Las subunidades alfa poseen sitios de unión para el potasio en su cara extracelular y para el sodio en su cara intracelular.
La bomba transporta ambos cationes en sentido opuesto (antiporte) y contra sus respectivos gradientes. El Na+ es transportado hacia el medio extracelular y el K+ es transportado hacia el medio intracelular. El transporte de Na+ y el de K+ están acoplados, pues no pueden realizarse el uno sin el otro.
El sistema funciona con aporte de energía, proporcionada por la hidrólisis de ATP. La subunidad alfa posee, en su cara citosólica, un sitio para el ATP y cataliza su hidrólisis.
Cada ATP que se hidroliza posibilita el transporte de 3 Na+ hacia el espacio extracelular y 2 K+ hacia el citosol.
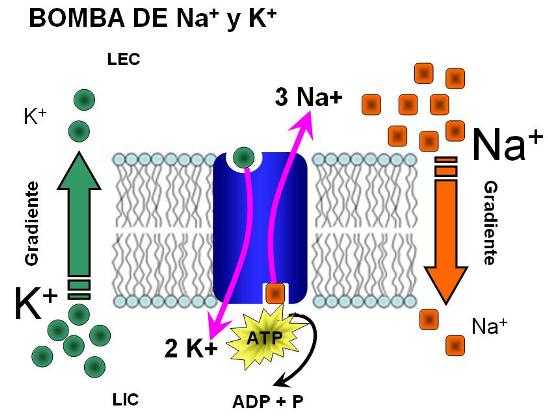 |
La bomba de Na+ y K+ es la responsable de la desigualdad en la distribución de estos cationes entre el LIC y el LEC.
La bomba de Na+ y K+ también es electrogénica: genera una diferencia de voltaje o potencial eléctrico a ambos lados de la membrana. El potencial eléctrico se establece cuando las cargas eléctricas a uno y otro lado no están equilibradas. Como la bomba extrae tres cationes por cada dos que introduce, contribuye a que la membrana plasmática sea más negativa en su cara citosólica que en su cara extracelular.
Transporte en masa Regresar
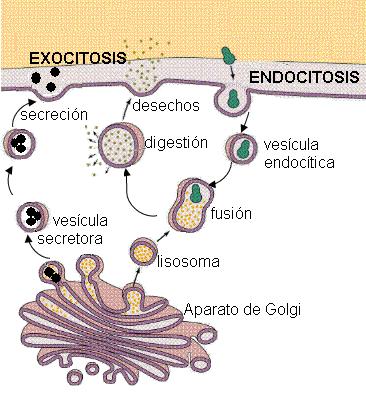
La membrana plasmática puede transportar macromoléculas, como proteínas, y también partículas de mayor tamaño. El transporte de este tipo de sustancias está mediado por vesículas y se denomina transporte en masa. La endocitosis y la exocitosis son dos formas del transporte en masa.
La endocitosis consiste en la incorporación de grandes moléculas que están en suspensión en el medio extracelular. En el sitio donde se produce el ingreso, la membrana se invagina, formando una depresión, de cuya parte más profunda se desprende hacia el citosol una vesícula endocítica o “endosoma”. En el endosoma quedan encerradas las macromoléculas. Posteriormente, el endosoma puede fusionarse con un lisosoma, lo que posibilita la digestión o hidrólisis de las sustancias incorporadas.
La exocitosis es un transporte por el cual las células pueden exportar productos que, provenientes del aparato de Golgi, fueron empacados en vesículas de secreción. Las vesículas de secreción se fusionan con la membrana plasmática y vuelcan su contenido al exterior. Así se liberan al medio extracelular hormonas, componentes de la matriz extracelular, enzimas, anticuerpos y muchas otras sustancias útiles fuera de la célula de origen.
En algunos casos la exocitosis permite la eliminación de desechos del proceso digestivo ocurrido dentro de los lisosomas.
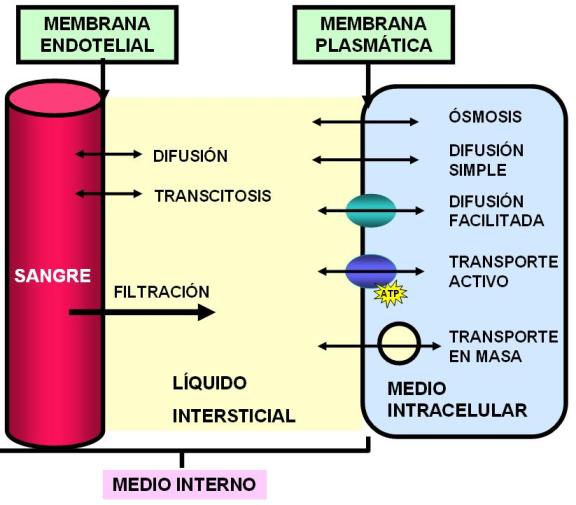
Intercambios a través de la membrana endotelial Regresar
Los dos líquidos corporales que forman el medio interno, el plasma y el líquido intersticial, están separados por la membrana endotelial.
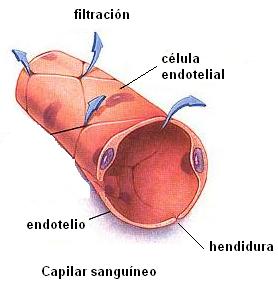
El endotelio es la capa de tejido interior en las paredes de los vasos sanguíneos. Las arterias y las venas tienen gruesas paredes con tejido muscular y conectivo por fuera del endotelio. Esto las convierte en estructuras impermeables. Los vasos sanguíneos capilares, en cambio, tienen una pared muy delgada formada solamente por la membrana endotelial. Son permeables, y por eso se constituyen en la superficie de intercambio entre el plasma y el líquido intersticial.
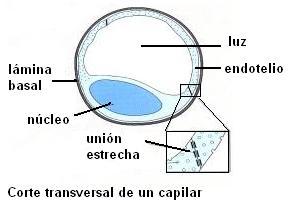 |
El endotelio es un epitelio formado por una sola capa de células planas, rodeadas por una membrana basal. Las células endoteliales están unidas mediante uniones estrechas separadas por poros o hendiduras. El diámetro de estas hendiduras es algo menor que el de un molécula proteica, como la albúmina.
Por lo tanto, a través de las hendiduras el líquido puede filtrarse libremente, junto con la mayor parte de los iones y pequeñas moléculas disueltas, a excepción de las proteínas. A medida que la sangre atraviesa el capilar, un número enorme de moléculas de agua y de partículas disueltas difunden en ambos sentidos a través de la pared capilar, proporcionando una mezcla continua entre el plasma y el líquido intersticial.
Las moléculas liposolubles directamente difunden a través de la membrana plasmática de las células endoteliales.
Algunas macromoléculas son transportadas por transcitosis: se endocitan en una de las superficies de la pared endotelial y se exocitan por la superficie opuesta.
Las proteínas plasmáticas, que se encuentran en suspensión coloidal, generan una presión osmótica que se opone a la fuerza de filtración capilar. Esta presión, llamada presión oncótica o coloidosmótica evita una pérdida significativa de volumen de líquido desde la sangre al espacio intersticial.
Resumen de los intercambios entre LIV, LIT y LIC |
 |
Composición de los líquidos corporales Regresar
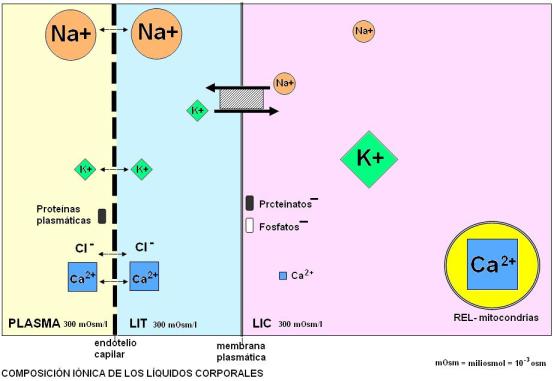
En el siguiente gráfico se observa la distribución en cada compartimiento de algunos iones de importancia fisiológica. Cada ión se representa con un símbolo diferente. Los tamaños de los respectivos símbolos intentan dar idea de la proporción relativa o concentración a que éstos se encuentran en los distintos líquidos corporales.
Como se puede observar, los iones inorgánicos sodio (Na+), potasio (K+), cloruro (Cl-) y calcio (Ca2+) se hallan a igual concentración en el líquido intersticial y en el plasma, ya que filtran libremente por los poros capilares. La principal diferencia entre estos compartimientos está dada por las proteínas plasmáticas, cuyo tamaño, superior al de los poros del endotelio, hace que sean retenidas en el plasma.
Cada compartimiento tiene neutralidad eléctrica: la suma de los aniones es igual a la de los cationes. La presencia de iones no difusibles a un lado de la membrana afecta la distribución de los iones difusibles. Por ejemplo, la carga negativa de las proteínas plasmáticas (no difusibles) obstaculiza la difusión de los cationes difusibles (que son atraídos) y favorece la de los aniones difusibles (los cuales resultan repelidos) desde la sangre.
El líquido intracelular contiene proteinatos (aniones de proteínas con cargas negativas) y fosfatos. Su concentración de cloruros es poco significativa. En lo que respecta al Na+ y al K+, sus proporciones se invierten en relación al plasma; mientras que el Na+, es el principal catión extracelular, el K+ es el principal catión intracelular. Esta diferencia es el resultado del continuo accionar de la bomba de y K+. En cuanto al anión cloruro, se acumula en el LEC pues se moviliza tras el Na+, siguiendo un gradiente eléctrico.
La concentración de Ca2+ es extremadamente baja en el citosol; en cambio es muy alta en el REL y las mitocondrias. Esto obedece a que en las membranas de estas organelas existen bombas de Ca2+ que lo acumulan contra gradiente, con gasto de ATP.
A pesar de las diferencias en la composición iónica, los líquidos en los tres compartimientos tienen la misma osmolaridad (concentración de partículas medida en osmoles / l) igual a 300 miliosmoles /l. Todos los líquidos corporales se encuentran en equilibrio osmótico.
El término tonicidad se usa para describir la osmolaridad de una solución comparada con el plasma. Si una solución tiene la misma osmolaridad que el plasma es isotónica; cuando tiene mayor osmolaridad es hipertónica; si su osmolaridad es menor que la del plasma, es hipotónica.
 |
|