|
Capítulo 12: Sistema Digestivo
Introducción Regresar
El acto de comer es una actividad que las personas, igual que el resto de los animales, realizamos cotidianamente. Los seres humanos comemos cuando tenemos hambre, pero también por placer, por costumbre, por festejar, porque estamos aburridos, por muchos motivos diferentes. No siempre somos concientes de que comemos para satisfacer una necesidad fisiológica ni de la importancia que tiene nuestra alimentación para el crecimiento, durante la niñez y la adolescencia, o para nuestro bienestar y salud, tanto actual como futura.
Pero... ¿Por qué necesitamos comer? El organismo está constituido por unidades llamadas células; el latido del corazón, correr, pensar, respirar, son el resultado de la actividad de millones de células. Para construirse a sí mismas y para funcionar, las células deben incorporar continuamente una cantidad de sustancias que no pueden sintetizar: los nutrientes. Los alimentos que componen la dieta son las fuentes de donde obtenemos los nutrientes.
El sistema digestivo es el encargado de la incorporación de los alimentos y de la transformación de los mismos, posibilitando que los nutrientes contenidos en aquéllos ingresen al medio interno. Una vez en el medio interno, los nutrientes están accesibles a las células, donde son asimilados.
El sistema digestivo también se ocupa de la eliminación de los componentes del alimento que no se incorporan al medio interno, junto con los cuales se desechan algunos residuos del proceso digestivo.
Anatomía del sistema digestivo Regresar
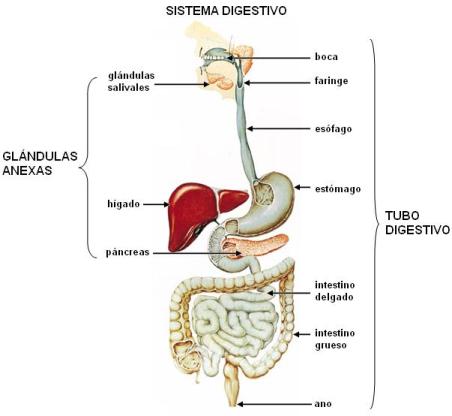
El sistema o aparato digestivo consta del tubo digestivo y las glándulas anexas.
El tubo digestivo se inicia en la boca y termina en el ano y en él se diferencia una serie de órganos que comparten una estructura común y a lo largo de los cuales el alimento ingerido avanza a medida que va siendo modificado.
Los órganos del tubo digestivo son:
· Cavidad bucal
· Faringe
· Esófago
· Estómago
· Intestino delgado
· Intestino grueso
· Conducto anal
Las glándulas anexas se originan durante el desarrollo embrionario como evaginaciones de las paredes del tubo y se mantienen conectadas al mismo por medio de sus conductos excretores. Comprenden:
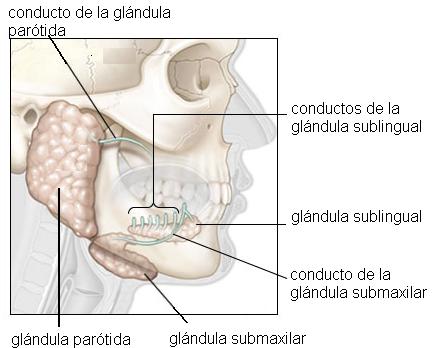
· Las glándulas salivales (glándulas pares) llamadas parótidas, submaxilares y sublinguales, que secretan la saliva hacia la cavidad bucal.
· El hígado, que secreta la bilis hacia el duodeno, primera porción del intestino delgado.
· El páncreas, que vuelca su secreción, el jugo pancreático, en el duodeno.
Histología del tubo digestivo Regresar
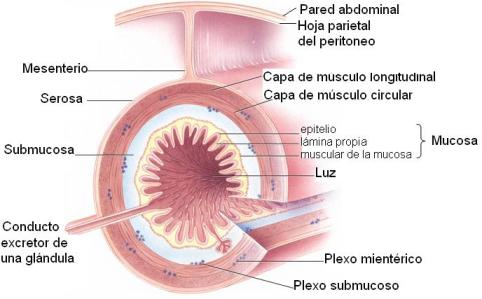
Los órganos que forman el tubo digestivo presentan una estructura histológica común, que consiste en cuatro capas de tejido. Desde la luz del tubo hacia la periferia, éstas son:
· Mucosa
· Submucosa
· Muscular
· Serosa
La mucosa es una capa de tejido epitelial en contacto con la luz de los órganos. El tejido epitelial se adapta a distintas funciones y presenta diferentes formas según el órgano. Se especializa en la protección, la secreción de moco, la secreción de enzimas y la absorción. Apoya sobre una lámina basal o lámina propia.
En el límite externo de la mucosa se halla la muscular de la mucosa (“muscularis mucosae”), formada por tejido muscular liso.
La submucosa está formada por tejido conectivo. Posee abundante irrigación e inervación. El tejido nervioso forma el plexo submucoso o plexo de Meissner. La submucosa está recorrida por glándulas que se invaginan desde la capa mucosa adyacente.
La muscular consta de tejido muscular liso dispuesto típicamente en dos capas: una interna de disposición circular y otra externa, de disposición longitudinal. Una variante se encuentra en el estómago, donde se agrega una capa de fibras musculares dispuestas en forma oblicua. También se diferencia el intestino grueso, en el cual la capa de músculo longitudinal es discontinua, formando bandas.
En la muscular se encuentra el plexo de Auerbach o mientérico (mio: músculo; enterón: intestino).
El tejido muscular liso de esta capa es responsable de la motilidad del tubo digestivo. Se halla inervado por el sistema nervioso autónomo simpático y parasimpático.
En el tercio superior del esófago las fibras musculares son estriadas.
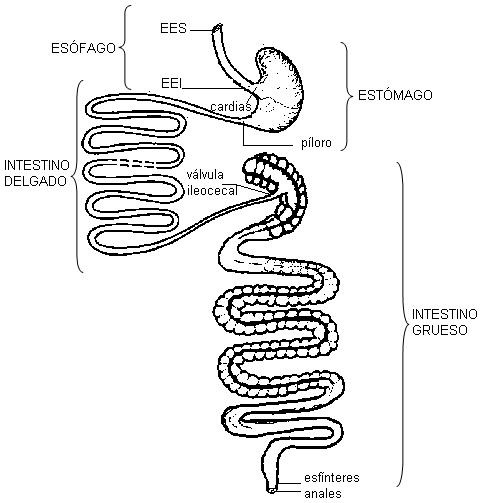
La muscular forma los esfínteres, que regulan el avance del alimento desde un órgano a otro:
· El esfínter esofágico superior (EES), en el inicio del esófago.
· El esfínter esofágico inferior (EEI), en el límite entre el esófago y el estómago.
· El píloro, ubicado entre el estómago y el duodeno.
· La válvula ileocecal, que separa al intestino delgado del ciego, la primera porción del intestino grueso.
· El esfínter anal. Éste tiene dos partes: el esfínter anal interno, formado por músculo liso, y el esfínter anal externo, formado por músculo estriado, de control voluntario.
La serosa está formada por tejido epitelial y reviste todos los órganos del tubo ubicados en la cavidad abdominal (estómago, intestino delgado e intestino grueso).
La capa serosa es una dependencia del peritoneo. Esta membrana posee dos hojas: una visceral, que envuelve a los órganos, y otra parietal, que se aplica contra las paredes internas de la cavidad abdominal. Los mesos son las porciones del peritoneo que conectan a los órganos con las paredes del abdomen. A través de los mesos los órganos reciben irrigación e inervación.
Entre las hojas visceral y parietal del peritoneo existe una cavidad virtual, la cavidad peritoneal.
Además del tubo digestivo, también el hígado y el bazo se hallan envueltos por el peritoneo.
En el esófago, que atraviesa el cuello y el tórax, la serosa está reemplazada por un tejido conectivo que forma la capa adventicia.
Fisiología general del aparato digestivo Regresar
El aparato digestivo, junto a los aparatos respiratorio y urinario, constituye una superficie de contacto e intercambio entre el medio externo y el medio interno.
La función primordial del aparato digestivo consiste en aprovisionar al medio interno de diferentes sustancias indispensables para la supervivencia: los nutrientes.
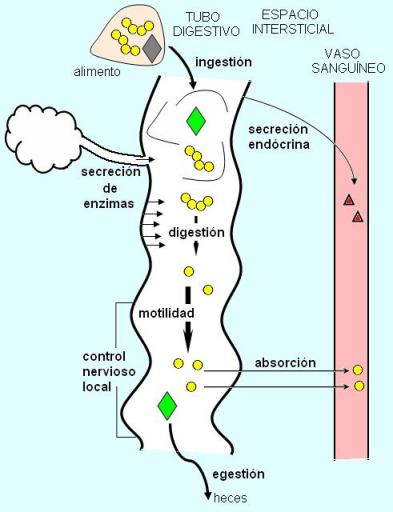
Para llevar a cabo dicha función, en el aparato digestivo tienen lugar diferentes procesos. El aparato digestivo recibe los alimentos cuando se produce la ingestión y los descompone hasta liberar los nutrientes que están contenidos en ellos. La descomposición o simplificación de la estructura del alimento recibe el nombre de digestión.
La digestión mecánica consiste en una simplificación que no afecta la estructura molecular. Incluye la humectación y la disgregación física de las partículas alimenticias. La masticación y los movimientos producidos por la capa muscular del tubo digestivo contribuyen a separar al alimento en partículas más pequeñas, lo que no solo facilita su avance a lo largo del tubo, sino que también lo prepara para una eficaz digestión química.
La digestión química es un proceso de hidrólisis, que divide a las moléculas del alimento en sus unidades constitutivas. Por ejemplo, separa a los aminoácidos de las cadenas proteicas o a los monosacáridos de una cadena de almidón.
La hidrólisis requiere la intervención de enzimas hidrolíticas específicas. Las enzimas digestivas se especializan en la hidrólisis de determinados enlaces (peptídico, glucosídico, éster, etc.) y están contenidas en los diversos jugos que actúan sobre el alimento en la luz del tubo digestivo. Dichas enzimas son secretadas por las glándulas anexas, como las salivales y el páncreas, o bien por células especializadas de la mucosa de los órganos que conforman el tubo, como la mucosa gástrica (gastros: estómago) y la del intestino delgado. El intestino delgado también posee enzimas que se hallan insertas en la membrana apical de las células de la mucosa.
Las enzimas digestivas actúan a diferentes valores de pH; los jugos digestivos contienen, además de enzimas, otras secreciones cuya función es generar las condiciones de acidez o alcalinidad necesarias para una actividad hidrolítica óptima en cada órgano.
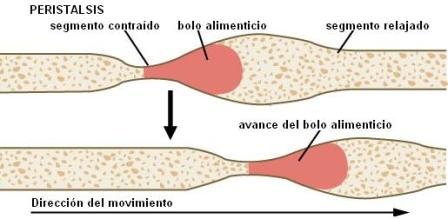
La musculatura del tubo digestivo lleva a cabo movimientos de mezcla y movimientos peristálticos. El peristaltismo propulsa los alimentos mediante la combinación de la contracción muscular de un área y la relajación de la siguiente. Una vez que el alimento avanza hacia la zona relajada, ésta se contrae, impulsándolo hacia adelante.
Los esfínteres impiden el reflujo del alimento y se abren oportunamente para permitir el avance desde un órgano hacia el siguiente.
Mientras los alimentos o los productos de su digestión permanecen en la luz del tubo digestivo, puede decirse que aún no han sido verdaderamente incorporados, pues no están accesibles a todas las células del cuerpo. La verdadera incorporación se produce cuando los productos de la digestión llegan al medio interno. Para ello, deben atravesar las paredes del tubo digestivo, que es una superficie de intercambio entre el medio externo (la luz del tubo digestivo) y el medio interno. Se denomina absorción al pasaje de los nutrientes (u otras sustancias) desde la luz del tubo digestivo al medio interno. Las sustancias absorbidas se incorporan generalmente a la circulación sanguínea o, en algunos casos, a los vasos linfáticos.
En la absorción influyen diversos factores; un factor fundamental es el tamaño de las partículas. La digestión, en tanto transforma a las macromoléculas en moléculas pequeñas, es un paso necesario para permitir la absorción. Salvo excepciones, como la absorción de alcohol o ciertos medicamentos, que puede realizarse en las paredes del estómago, la absorción tiene lugar mayoritariamente en el intestino delgado y, en menor medida, en el intestino grueso.
Es importante destacar que la absorción es selectiva en el sentido en que puede serlo una membrana, es decir dependiendo del tamaño de las partículas, de sus gradientes y del tipo de transportadores presentes, y no en un sentido “inteligente”. Las membranas no discriminan si las sustancias absorbidas serán útiles o perjudiciales.
Además de nutrientes, en el intestino se absorbe gran parte del agua, los electrólitos y otros componentes de los jugos digestivos, como las sales biliares.
Las sustancias del alimento que no pueden ser digeridas y/o absorbidas se expulsan en la materia fecal. Ésta contiene también ciertas excreciones o desechos, como los pigmentos biliares. La expulsión de la materia fecal se denomina egestión.
Los alimentos que ingresan al aparato digestivo no son estériles, por el contrario, tienen bacterias y otros elementos que pueden resultar nocivos. Gran parte de las bacterias mueren debido a la acidez del jugo gástrico. Además, las paredes intestinales presentan grandes acumulaciones de tejido linfático, las placas de Peyer, que ejercen protección contra agentes invasores.
El sistema digestivo está controlado por el sistema nervioso autónomo y muchas de sus funciones se regulan mediante reflejos locales, integrados en los plexos nerviosos de sus propias paredes (el “sistema nervioso entérico”).
En las paredes del tubo digestivo hay células endócrinas especializadas que secretan hormonas (principalmente péptidos) algunas de la cuales tienen acción local, mientras que otras producen sus efectos a nivel sistémico.
En conclusión, el aparato digestivo cumple múltiples funciones:
· Ingestión
· Motilidad
· Secreción
· Digestión mecánica
· Digestión química
· Absorción
· Egestión
· Protección inmunológica
· Control nervioso y endócrino
Nutrientes Regresar
Los nutrientes pueden clasificarse en cinco grupos: glúcidos, lípidos, proteínas, vitaminas y minerales. Los glúcidos y los lípidos cumplen una función eminentemente energética; las proteínas desempeñan una función plástica o reparadora, pues se utilizan como “materiales de construcción” en las células; las vitaminas y los minerales ejercen funciones reguladoras.
Los glúcidos, lípidos y proteínas son los nutrientes cuantitativamente más importantes (macronutrientes), en tanto las vitaminas y los minerales son indispensables en muy pequeñas cantidades (micronutrientes).
Los glúcidos de la dieta están representados principalmente por almidón, disacáridos y monosacáridos. Los glúcidos se digieren hasta monosacáridos y son absorbidos como tales.
Otros polisacáridos diferentes del almidón, como la celulosa, son indigeribles y forman parte de la llamada “fibra alimentaria”.
Desde el punto de vista alimentario, los lípidos más importantes son los triacilglicéridos (aceites y grasas) y el colesterol. Los primeros son hidrolizados a glicerol, monoacilglicéridos y ácidos grasos libres, antes de ser absorbidos. El colesterol se absorbe como tal; si está combinado con ácidos grasos, previamente a la absorción se hidroliza la unión entre ambos.
Es importante que los lípidos de la dieta incluyan a los ácidos grasos linoleico y linolénico, de las series n-6 y n-3, respectivamente, pues éstos son esenciales, es decir que no pueden ser sintetizados en el organismo por transformación de otros ácidos grasos.
Los aceites o “grasa insaturada”, de origen vegetal, deben predominar en la dieta sobre las “grasas saturadas”, de origen animal, ya que estas últimas, al igual que las grasas “trans” son aterogénicas.
Las proteínas alimentarias son la fuente de aminoácidos para la síntesis de proteínas. Si bien muchos aminoácidos pueden ser sintetizados en las células, los aminoácidos esenciales deben estar presentes en las proteínas ingeridas. Las proteínas de origen animal son consideradas “completas”, pues aportan aminoácidos esenciales en mayores y más adecuadas proporciones que las proteínas de origen vegetal, nutricionalmente “incompletas”.
Las proteínas se hidrolizan hasta cadenas cortas de aminoácidos (dipéptidos y tripéptidos) y aminoácidos libres, lo que posibilita su absorción.
Las vitaminas y minerales generalmente se absorben con muy poca o ninguna transformación en el aparato digestivo.
Entre las vitaminas se hallan las hidrosolubles, que comprenden a los distintos compuestos del “complejo B” y la vitamina C (ácido ascórbico), y las liposolubles, que incluyen a las vitaminas A, D, E y K. La absorción y posterior metabolismo de estas últimas están ligados a los de las grasas.
Nutrientes |
Macronutrientes |
Glúcidos |
Energéticos |
Lípidos |
Proteínas |
Reparadores |
Micronutrientes |
Vitaminas |
Reguladores |
Minerales |
Boca: digestión bucal Regresar
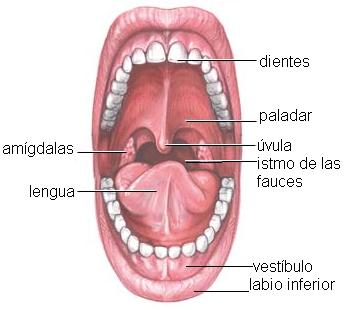
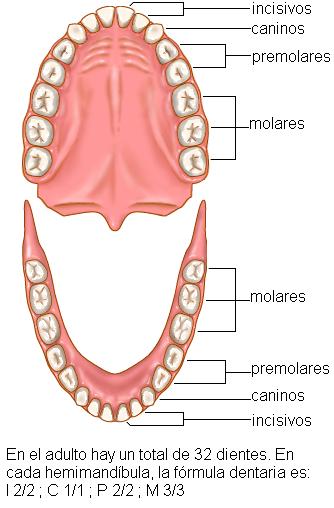
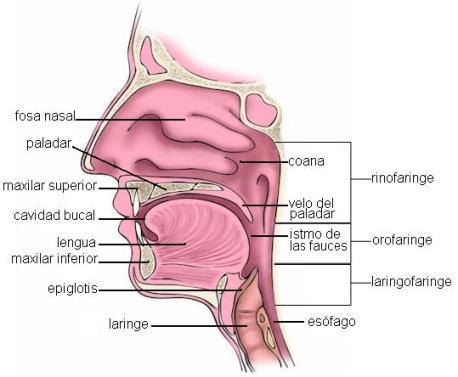
La cavidad bucal se describe como una cavidad con seis caras: un piso, un techo, una cara anterior, otra posterior y dos paredes laterales. En la cara anterior se encuentra el orificio bucal, limitado por los labios superior e inferior; entre éstos y las arcadas dentarias se ubica el vestíbulo bucal. En el piso de la boca se encuentra la lengua y el techo corresponde al paladar, que separa la cavidad bucal de las fosas nasales. Las caras internas de las mejillas forman las paredes laterales. La pared posterior está flanqueada por los pilares del paladar y conduce a la faringe a través del istmo de las fauces.
 |
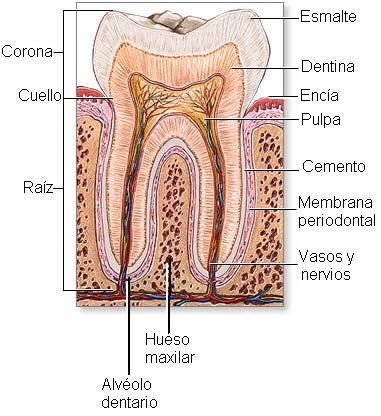 |
Cada diente posee una corona, un cuello y una raíz, la cual está inserta en el alvéolo dentario, cavidad excavada en el hueso maxilar.
De afuera hacia adentro, un diente presenta tres capas: el esmalte, la dentina o marfil y la pulpa. En esta última se ubican los vasos y nervios del diente. |
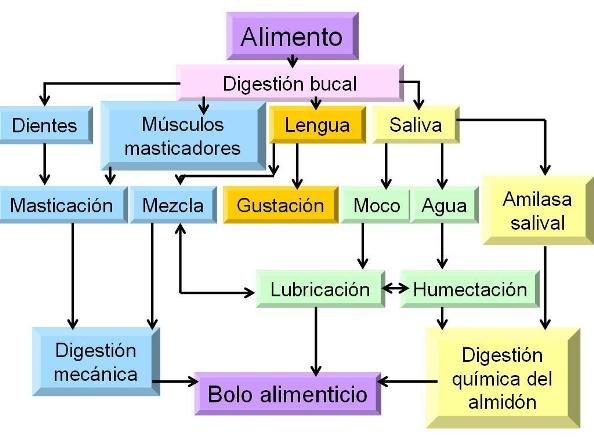
En la cavidad bucal tienen lugar los primeros procesos de digestión mecánica y química. La digestión mecánica es el resultado de la masticación. La digestión química se debe a una enzima presente en la saliva, la ptialina o amilasa salival, que actúa sobre las moléculas de almidón, produciendo la hidrólisis parcial de las mismas. De dicha hidrólisis se obtienen moléculas de maltosa, pocas unidades de glucosa libre y dextrinas. Estas últimas son polímeros cortos de glucosa que contienen los puntos de ramificación, los cuales no son atacados por la ptialina. La ptialina actúa en medio alcalino.
La saliva no solo participa en la digestión química, sino que además es indispensable para la masticación y la aglutinación de las partículas alimenticias. El mucus contenido en la saliva lubrica el alimento y contribuye a transformarlo en un bolo alimenticio.
 |
La lengua ayuda a la formación del bolo alimenticio y posee las papilas gustativas, donde se encuentran los receptores del sentido del gusto.
Una vez formado el bolo alimenticio, los movimientos de la lengua lo empujan hacia el istmo de las fauces, iniciando el proceso de deglución.
Faringe y esófago: Deglución Regresar
La deglución es el pasaje del bolo alimenticio desde la boca hasta el estómago. Durante la deglución el bolo alimenticio recorre la faringe y el esófago.
La faringe es un órgano común a los aparatos digestivo y respiratorio; da paso al bolo alimenticio y al aire durante la inspiración y la espiración.
La faringe es un tubo de paredes musculares, de unos 13 cm de largo por 3 cm de diámetro. Este tubo está cerrado en su parte posterior, pero por su parte anterior comunica sucesivamente con las fosas nasales (a través de las coanas), con la boca (por medio del istmo de las fauces) y con la laringe. Así quedan delimitadas sus tres porciones: la rinofaringe, la orofaringe y la laringofaringe.
Hacia abajo la faringe se continúa con el esófago, ubicado por detrás de la laringe.
El esófago es un conducto que ocupa sucesivamente el cuello y el tórax, y atraviesa el diafragma ingresando en el abdomen, donde se continúa con el estómago. El tejido muscular forma los esfínteres esofágicos superior e inferior en ambos extremos del conducto, los cuales regulan el ingreso del bolo al esófago y su pasaje hacia el estómago, respectivamente.
 |
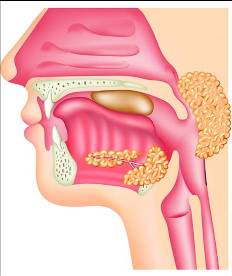
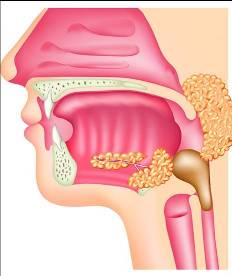
La faringe y el esófago participan en la deglución. La deglución se inicia voluntariamente cuando el bolo alimenticio es empujado por la lengua hacia la faringe. A partir de la estimulación de la faringe por el bolo alimenticio, la deglución se convierte en un acto reflejo (involuntario). Se desencadena entonces una serie de mecanismos que ayudan al avance del bolo alimenticio por el tubo digestivo. Al mismo tiempo, cierran la vía respiratoria para evitar que el bolo se desvíe hacia la misma. Por lo tanto, la respiración queda interrumpida durante el instante en que ocurre la deglución.
Los mecanismos reflejos de la deglución incluyen:
- La elevación del velo del paladar, que cierra las coanas.
- El ascenso de la laringe y el cierre de su orificio superior, la glotis, que queda cubierta por un cartílago a modo de tapa (la epiglotis).
- La contracción de los músculos faríngeos, que empujan el bolo hacia abajo.
- La relajación del esfinter esofágico superior, para permitir el ingreso del bolo alimenticio.
 |
 |
Una vez en el esófago, el bolo avanza gracias a los movimientos peristálticos, ondas de contracción que recorren el órgano en sentido descendente. La llegada de las ondas peristálticas a la zona del EEI ocasiona la relajación del mismo, permitiendo el ingreso del bolo alimenticio al estómago.
En su paso por la faringe y el esófago el bolo alimenticio no sufre modificaciones.
El epitelio que constituye la mucosa de ambos órganos está adaptado para resistir agresiones físicas, pues se trata de un epitelio pluriestratificado plano. Además, posee glándulas mucosas, cuyas secreciones lubrican el conducto facilitando el avance del alimento.
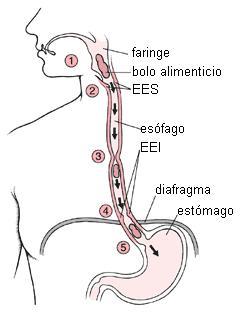 |
1- El bolo alimenticio es empujado hacia la faringe.
2- Se relaja el ESS.
3- Las ondas peristálticas propulsan al bolo alimenticio.
4- Se relaja el EEI.
5- El bolo alimenticio ingresa al estómago. |
Estómago: Digestión gástrica Regresar
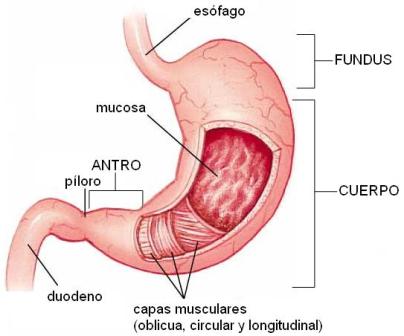 |
El estómago (gastros) es una porción dilatada del tubo digestivo, cuya forma se compara con una gaita o una letra J mayúscula. Se encuentra entre el esófago y el duodeno, con los cuales se comunica a través de sendos orificios: el cardias y el píloro. Desde el punto de vista anatómico se divide en tres regiones: el fundus, el cuerpo y el antro.
El estómago se comporta como un reservorio, pues adapta su capacidad (de aproximadamente 1500cc) para recibir distintas cantidades de alimento.
En el estómago, el bolo alimenticio se mezcla con el jugo gástrico hasta convertirse en una pasta llamada quimo. En la formación del quimo se combinan fenómenos mecánicos y fenómenos químicos.
La digestión mecánica consiste en movimientos de mezcla que diluyen el bolo alimenticio en el jugo gástrico y trituran las partículas sólidas de alimento hasta que alcanzan un tamaño muy pequeño.
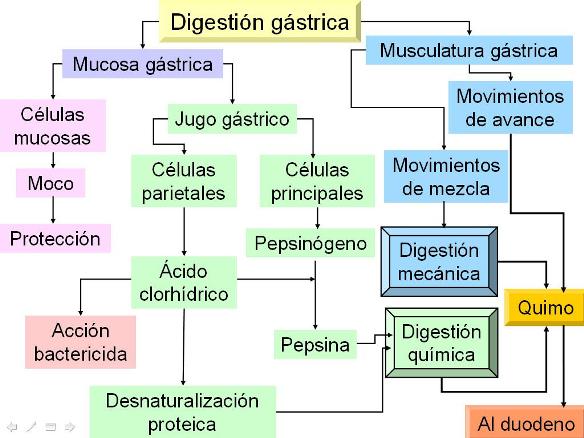
En la digestión química participa el jugo gástrico, secretado por la mucosa gástrica. Ésta consta de un epitelio cilíndrico simple, con células productoras de moco y numerosas glándulas tubulares, llamadas “criptas”.
La composición del jugo gástrico es diferente cuando el estómago está en reposo o en actividad. El jugo gástrico en reposo es secretado por las células superficiales; tiene una composición similar al plasma, con mayor concentración de bicarbonato (HCO3-), que actúa como base, y pH alcalino. Las células mucosas superficiales también secretan una gruesa capa de moco (una glucoproteína) el cual protege a la mucosa de la acidez del jugo gástrico, cuando el estómago se halla en actividad.
Fallas en la barrera mucosa del estómago pueden provocar una lesión de sus paredes, la úlcera péptica.
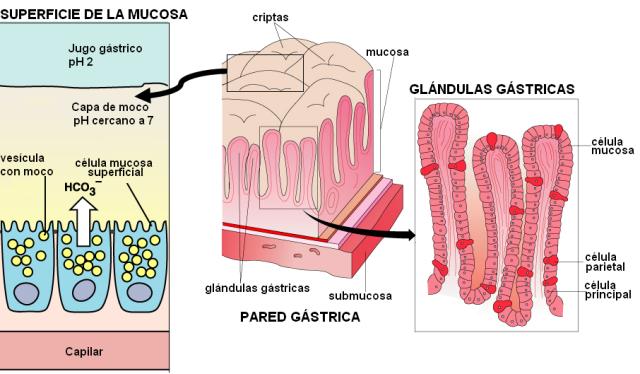 |
En las glándulas gástricas se encuentran diferentes tipos celulares:
• Células mucosas: se ubican en el cuello de las glándulas. Contienen REG y aparato de Golgi bien desarrollados y secretan un mucus alcalino.
• Células parietales u oxínticas: poseen abundantes mitocondrias; son las encargadas de la secreción de ácido clorhídrico y del factor intrínseco (proteína que se une a la vitamina B12 y permite su absorción en el intestino delgado).
• Células principales o zimógenas: son típicas células secretoras de proteínas; secretan el pepsinógeno.
• Células indiferenciadas: se ubican en el cuello de la glándula y se reproducen, permitiendo la renovación de la mucosa.
• Células APUD (sigla de “captación y decarboxilación de precursores de aminas”): son células con función endócrina.
Cuando el estómago es estimulado, ya sea por la visión u olfacción de los alimentos, o por la llegada del bolo alimenticio, produce un jugo gástrico ácido, cuyo pH es cercano a 2, debido a la secreción de ácido clorhídrico (HCl).
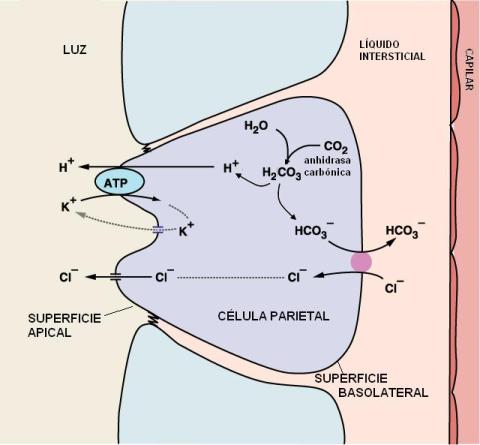
Las células parietales poseen una membrana apical muy plegada, formando canalículos, donde se encuentran los transportadores que participan en la secreción del ácido clorhídrico.
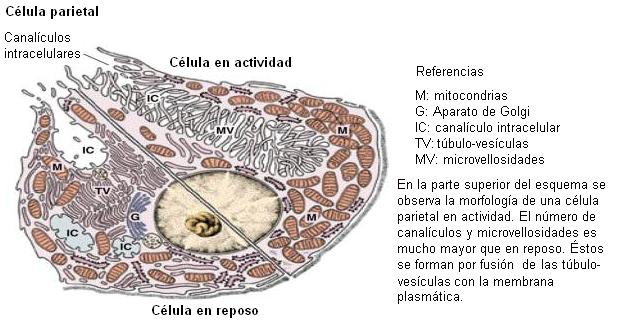 |
Los componentes del ácido clorhídrico, protones (H+) y cloruro (Cl-), se secretan por separado.
Los H+ son movilizados hacia el lumen mediante un tipo de transporte activo primario, a cargo de una bomba H+/K+ ATPasa. La bomba transporta H+ y K+ en sentido opuesto, ambos contra sus respectivos gradientes de concentración, con gasto de ATP.
Los protones se generan en la célula parietal por disociación del ácido carbónico (H2CO3), el cual es el producto de una reacción entre agua y CO2, catalizada por la enzima anhidrasa carbónica. La disociación del ácido carbónico también produce bicarbonato (HCO3-). Éste es intercambiado por cloruro en la membrana basolateral de la célula parietal, por medio de un cotransportador. La secreción de bicarbonato es causante de la alcalinización de la sangre después de la ingesta, fenómeno conocido como “marea alcalina”.
El Cl- y el K+ salen hacia la luz del estómago por la membrana apical, a través de sendos canales y a favor de sus respectivos gradientes.
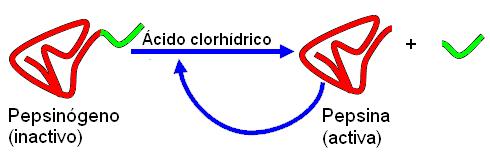
El ácido clorhídrico, además de actuar como un potente bactericida, es necesario en el jugo gástrico para la conversión del pepsinógeno en pepsina.
 |
Otro componente del jugo gástrico en la etapa de actividad es el pepsinógeno, un zimógeno secretado por las células principales. Los zimógenos son moléculas enzimáticas inactivas, hasta que pierden una parte de su cadena; entonces adquieren su actividad catalítica. La forma activa del pepsinógeno es la pepsina. La conversión del pepsinógeno a pepsina ocurre en el lumen gástrico, en contacto con el ácido clorhídrico. Las moléculas de pepsina son autocatalíticas, pues una vez formadas contribuyen a la activación de otras moléculas de pepsinógeno.
 |
La pepsina es una endopeptidasa, enzima que actúa sobre proteínas y polipéptidos, hidrolizando los enlaces peptídicos adyacentes a un tipo específico de aminoácidos, en el interior de la cadena. De esta forma, las cadenas proteicas y polipeptídicas quedan reducidas a moléculas más cortas, de diversos tamaños.
El jugo gástrico contiene también una lipasa (lipasa gástrica), enzima que hidroliza triacilglicéridos de cadena corta, aunque su actividad sólo cobra importancia en casos de insuficiencia pancreática.
Una vez que se ha completado la digestión gástrica, el píloro se relaja y movimientos de avance producidos por los músculos del estómago permiten evacuar su contenido, enviando el quimo hacia el duodeno.
Hígado: Secreción y acción de la bilis Regresar
El hígado es el órgano más grande del cuerpo y es vital. Se ubica en la parte superior del abdomen, debajo del diafragma, encima del estómago y de la masa intestinal.
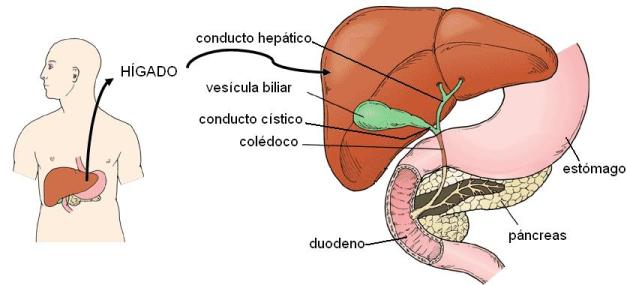 |
Las células hepáticas (hepatocitos) secretan la bilis en forma continua, la cual drena por la vía biliar intrahepática hasta el conducto hepático común. Desde allí, la bilis fluye hacia el conducto cístico y se acumula en la vesícula biliar, un receptáculo ubicado en la cara inferior del hígado. La vesícula biliar es un reservorio de bilis, que se contrae ante diversos estímulos, durante y después de las comidas, expulsando la bilis hacia el duodeno. La bilis llega al duodeno en forma intermitente, a través del conducto cístico y del colédoco, continuación del conducto hepático. El colédoco termina en la ampolla de Vater, junto con el conducto pancreático principal. La ampolla de Vater posee un esfínter, el esfínter de Oddi, y desemboca en un relieve de la pared duodenal, la carúncula mayor.
La bilis es un líquido amarillo-verdoso que contiene agua, electrólitos, sales biliares, colesterol, fosfolípidos y los pigmentos bilirrubina y biliverdina. Estos últimos, que le otorgan su color, son productos de desecho provenientes del metabolismo del grupo hemo.
La bilis se concentra en la vesícula biliar, ya que en las paredes de la misma se produce una reabsorción activa de Na+ que va acompañada de una reabsorción osmótica de agua.
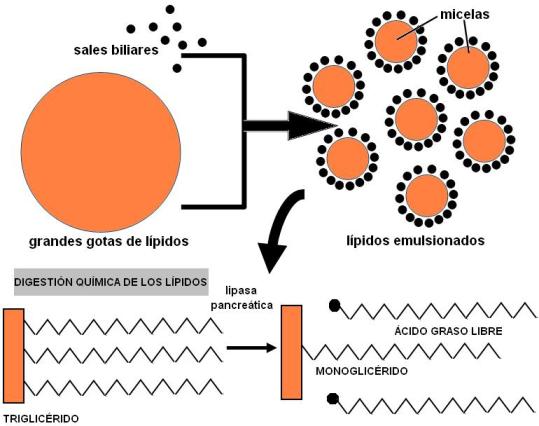
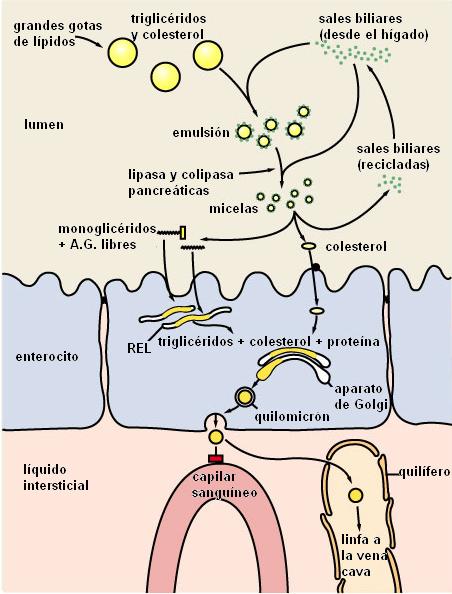
Las sales biliares son el componente activo de la bilis en el proceso digestivo; son sintetizadas en el hígado a partir del colesterol. Las sales biliares son detergentes, moléculas con una parte hidrofílica y otra parte hidrofóbica (anfipáticas), que actúan como emulsionantes de los lípidos de la dieta. Las sales biliares rompen las grandes gotas de lípidos (insolubles en agua) en gotas más pequeñas o micelas; éstas se mantienen suspendidas en el medio acuoso de los jugos digestivos, formando una emulsión. La emulsión de los lípidos es necesaria para la acción de las lipasas, enzimas que actúan en la digestión química de los lípidos.
Las sales biliares también se combinan con los productos de la digestión de los lípidos, aumentando su absorción a nivel intestinal. El déficit de sales biliares ocasiona una malabsorción de grasas.
Se requiere una pequeña cantidad de sales biliares, ya que la mayor parte de las mismas es reabsorbida en el íleon (la última porción del intestino delgado) y reciclada en el hígado.
La litiasis biliar es una enfermedad causada por la formación de piedras o cálculos (litos) por precipitación de los componentes de la bilis. Si se extirpa la vesícula biliar (colecistectomía), la bilis se secreta de forma constante y lenta hacia el duodeno, permitiendo la digestión de cantidades moderadas de grasas en la dieta.
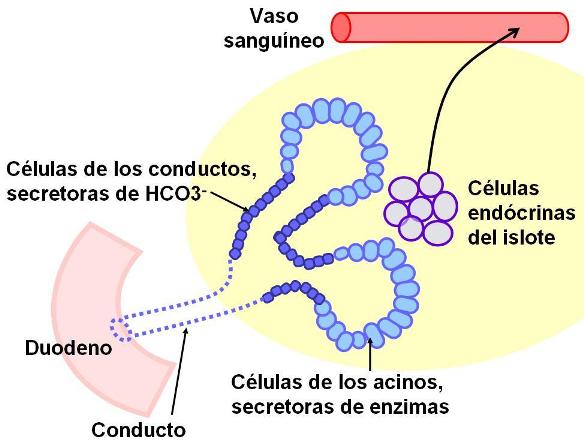
Páncreas y jugo pancreático Regresar
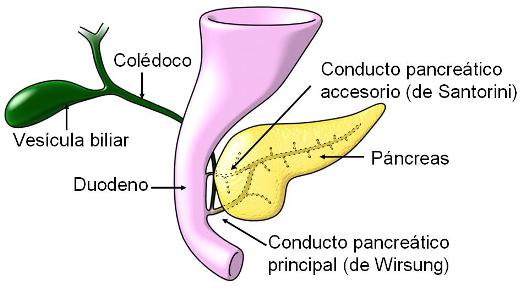
El páncreas es un órgano retroperitoneal, cuya cabeza se ubica en la curva que describe el duodeno. Los conductos que drenan su secreción exócrina, el jugo pancreático, son dos: el principal (conducto de Wirsung) y el accesorio (conducto de Santorini). El conducto de Wirsung desemboca en la carúncula mayor del duodeno, junto al colédoco. El conducto de Santorini termina en la carúncula menor, por encima del conducto de Wirsung.
 |
El jugo pancreático contiene sodio y bicarbonato, iones que, junto con la bilis y el jugo intestinal, neutralizan el quimo ácido proveniente del estómago, llevando el pH intestinal a 6 ó 7.
La secreción alcalina del páncreas no solo protege a la mucosa duodenal de la acidez del quimo, sino que también brinda un pH óptimo para la actividad de las enzimas pancreáticas.
El jugo pancreático es rico en enzimas digestivas que actúan sobre todos los macronutrientes: los hidratos de carbono, las grasas y las proteínas. Por esta razón, la digestión no se completa si la secreción de enzimas pancreáticas hacia el intestino delgado es defectuosa.
Cuando la aptitud de la glándula para sintetizar o secretar las enzimas se ve comprometida, se afectan la digestión y absorción de distintos componentes de la dieta. En estas circunstancias aumenta la cantidad de hidratos de carbono y proteínas en las heces. Sin embargo, el efecto predominante de la deficiencia de enzimas pancreáticas es la esteatorrea, o sea la aparición de grandes cantidades de grasa sin digerir en la materia fecal.
 |
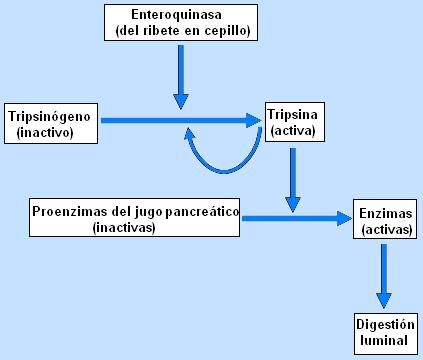
Varias de las enzimas pancreáticas se secretan en forma inactiva. Por ejemplo, las enzimas proteolíticas son secretadas como zimógenos o proenzimas y se activan en el lumen intestinal; de esta forma la glándula se protege de la autodigestión.
La tripsina es la enzima pancreática que activa a las demás proenzimas. Para impedir que la tripsina (y secundariamente las otras enzimas pancreáticas) se active dentro de la glándula, las células secretoras de enzimas de los acinos producen un inhibidor de la tripsina.
Obstrucciones de los conductos o lesiones en la glándula pueden provocar una acumulación de gran cantidad de secreción. En estas circunstancias, el inhibidor de la tripsina puede ser contrarrestado y las enzimas activadas. Se produce entonces un cuadro de pancreatitis aguda, que puede causar la muerte o una insuficiencia pancreática.
Las enzimas pancreáticas y sus respectivas acciones se resumen en el siguiente cuadro:
| Enzima |
Acción |
| Amilasa pancreática |
Hidroliza los enlaces interiores de las cadenas principales de la amilosa y la amilopectina. No puede hidrolizar los enlaces en los puntos de ramificación. |
| Lipasa pancreática |
Hidroliza los enlaces éster entre el glicerol y los ácidos grasos unidos a los carbonos 1 y 3. Los productos son monoacilglicéridos y ácidos grasos libres. |
| Procolipasa (zimógeno) -> Colipasa (activa) |
Activa a la lipasa. |
| Profosfolipasa A 2 (zimógeno) Fosfolipasa A 2 (activa) |
Actúa sobre los fosfoacilglicéridos, hidrolizando el enlace éster entre el glicerol y el ácido graso en posición 2. |
| Hidrolasa de ésteres de colesteril |
Hidroliza los ésteres de colesterol, liberando colesterol y un ácido graso. |
| Tripsinógeno (zimógeno) -> Tripsina (activa) |
Son endopeptidasas: hidrolizan enlaces peptídicos interiores a una cadena de aminoácidos. Cada una reconoce enlaces adyacentes a aminoácidos específicos. Liberan cadenas cortas de aminoácidos. |
| Quimotripsinógeno (zimógeno) -> Quimotripsina (activa) |
| Proelastasa (zimógeno) -> Elastasa (activa) |
Procarboxipeptidasa A (zimógeno)-> Carboxipeptidasa A (activa) |
Es una exopeptidasa: libera aminoácidos ácidos del extremo carboxiterminal de una cadena. |
| Procarboxipeptidasa B (zimógeno) -> Carboxipeptidasa B (activa) |
Es una exopeptidasa: libera aminoácidos básicos del extremo carboxiterminal de una cadena. |
| Ribonucleasa |
Hidroliza el ARN a nucleótidos. |
| Desoxirribonucleasa |
Hidroliza el ADN a nucleótidos. |
Estructura del intestino delgado Regresar
El intestino delgado, que se extiende desde el estómago hasta el colon, mide entre 6 y 8 metros y se divide en dos porciones: el duodeno y el yeyunoíleon. El duodeno es la primera porción del intestino delgado; mide unos 25 cm de longitud y está fijo a la pared posterior del abdomen mediante el peritoneo. El yeyunoíleon, la porción más larga del intestino delgado, es flotante; está sujeto a la pared posterior del abdomen por el mesenterio (meso del intestino), pero su borde anterior es libre. Ocupa la mayor parte del abdomen inferior, donde describe una serie de asas.
En el intestino delgado se lleva a cabo la fase más importante de la digestión. La hidrólisis de las sustancias orgánicas del alimento se completa en este órgano, de manera que los nutrientes quedan en condiciones de ser absorbidos. La mayor parte de la absorción también se produce en el intestino delgado.
La mucosa del intestino delgado está adaptada a funciones secretorias, digestivas y absortivas.
En la mucosa intestinal se encuentran dos tipos de glándulas: las criptas intestinales o glándulas de Lieberkühn y las glándulas de Brunner.
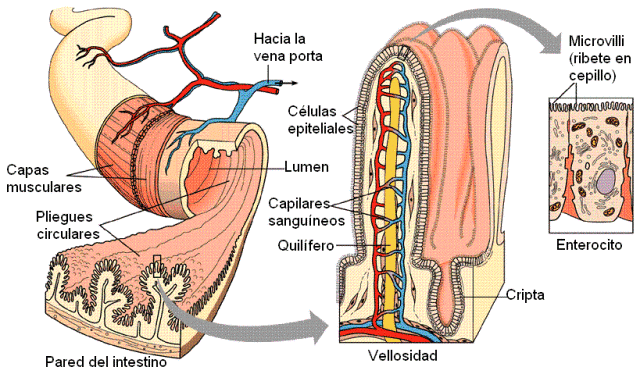
Las criptas intestinales son glándulas tubulosas que se extienden dentro de la capa mucosa, hasta la muscularis mucosae y se encuentran a lo largo de todo el intestino. Su desembocadura se ubica entre las vellosidades, que son elevaciones de la mucosa. Tanto las criptas como las vellosidades están tapizadas por un epitelio columnar simple formado por los enterocitos, en el que se intercalan células caliciformes, especializadas en la secreción de moco.
 |
Las células de la zona subterminal de las criptas intestinales sufren divisiones mitóticas y se diferencian dentro de la cripta, a medida que ascienden por la misma. De esta forma van “empujando” y reemplazando a los enterocitos que se descaman en el extremo distal de la vellosidad. El ciclo vital de una célula intestinal es de unos 5 días.
La cripta tiene una función predominantemente secretoria. Las secreciones de la cripta contienen agua y electrólitos que neutralizan la acidez del quimo recibido desde el estómago y contribuyen a mantener la isotonía del contenido luminal con el plasma.
La secreción acuosa de las criptas es reabsorbida luego en las vellosidades y, en condiciones normales, la absorción supera a la secreción.
En las criptas también se diferencian células con función endócrina y del sistema inmunitario.
Las glándulas de Brunner se encuentran en el duodeno; son túbulo-acinosas ramificadas y llegan hasta la submucosa. Son glándulas secretoras de moco. La función del moco secretado por las glándulas de Brunner consiste en proteger a la pared intestinal de la digestión por el jugo gástrico, pues el duodeno es un frecuente lugar de asiento de úlceras pépticas. La secreción de moco se activa ante diferentes estímulos irritantes.
La mucosa del intestino delgado presenta una serie de pliegues que hacen de ella una estructura altamente especializada para la función absortiva:
• En primer lugar, las válvulas conniventes o de Kerckring, pliegues circulares que triplican la superficie de absorción y se desarrollan principalmente en duodeno y yeyuno.
• En segundo lugar, las vellosidades, proyecciones digitiformes de alrededor de un milímetro de longitud, que sobresalen desde las válvulas conniventes. Éstas aumentan la superficie otras veinte veces.
• Por último, cada enterocito posee el ribete en cepillo, formado por las microvellosidades (microvilli). La superficie apical de un enterocito exhibe alrededor de mil microvellosidades, que incrementan el área superficial unas quince veces.
Se calcula que la superficie total de la mucosa intestinal, gracias a dichas estructuras, es equivalente a la de una cancha de tenis. Esta amplísima superficie optimiza la posibilidad de absorber los nutrientes que se liberan en la luz del intestino a medida que se lleva a cabo la digestión química.
La sustancias absorbidas generalmente deben atravesar el enterocito para llegar hasta la circulación, pues la vía paracelular se encuentra ocluida por las uniones estrechas.
La membrana plasmática de los enterocitos posee una superficie apical, en contacto con la luz, por donde ingresan los nutrientes. Éstos abandonan el enterocito a través de la superficie basal, en contacto con vasos sanguíneos y linfáticos. En ambas superficies, apical y basal, se ubican diferentes transportadores a través de los cuales se realiza la absorción.
En el interior de cada vellosidad se encuentran capilares sanguíneos y linfáticos, hacia donde se dirigen los nutrientes.
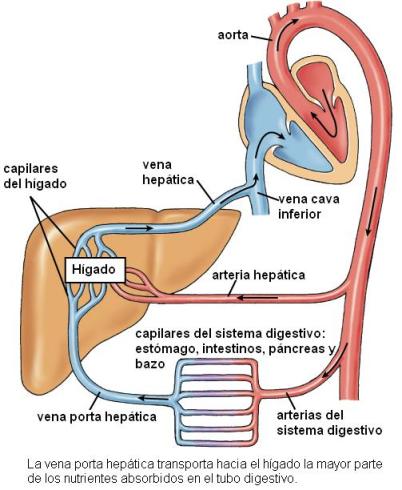
Los capilares sanguíneos pertenecen al sistema de la arteria mesentérica superior y confluyen luego en la vena mesentérica superior, tributaria de la vena porta. La vena porta transporta hasta el hígado los nutrientes absorbidos en las microvellosidades.
Los quilíferos son los capilares linfáticos que ocupan el centro de las microvellosidades. Hacia ellos se dirige la mayor parte de los lípidos obtenidos de la dieta.
 |
Digestión en el intestino delgado Regresar
En el intestino delgado hay procesos de digestión química y de digestión mecánica.
El intestino delgado recibe el quimo. Al duodeno se vuelcan también la bilis y el jugo pancreático, los cuales actúan sobre el quimo en la luz del intestino. Esta fase de la digestión química intestinal, que transcurre en la luz del órgano, se conoce como digestión luminal.
En el duodeno se completa la digestión química de los glúcidos y de las proteínas, iniciada en la boca y el estómago, respectivamente. A ello contribuyen las diversas enzimas del jugo pancreático. También se lleva a cabo la digestión de las grasas, gracias a la acción emulsificante de las sales biliares y la actividad hidrolítica de la lipasa pancreática.
Las células de la mucosa intestinal, los enterocitos, presentan una superficie apical plegada, formando microvellosidades; a esta superficie se la conoce como “el ribete en cepillo”. Las membranas del ribete en cepillo poseen diversas enzimas que participan en la digestión química. Una de estas enzimas es la enteroquinasa, que posee una unión débil a la membrana apical, por lo cual se libera fácilmente hacia el lumen. La enteroquinasa es la encargada de activar al tripsinógeno del jugo pancreático. La tripsina obtenida cataliza la activación de más moléculas de tripsinógeno y activa a su vez al resto de las proenzimas del jugo pancreático.
 |
En el ribete en cepillo se encuentran además otras enzimas hidrolíticas que actúan sobre los productos de la digestión luminal. La hidrólisis a cargo de las enzimas del ribete en cepillo es una digestión de superficie.
Las enzimas del ribete en cepillo son:
• Dextrinasas: hidrolizan dextrinas a monosacáridos.
• Disacaridasas: separan el disacárido en sus monosacáridos constitutivos.
• Aminopeptidasas: separan el aminoácido aminoterminal del péptido.
• Dipeptidasas: rompen los dipéptidos, liberando aminoácidos.
Como resultado de la acción de estas enzimas, los glúcidos de la dieta se reducen a monosacáridos y las proteínas a tripéptidos, dipéptidos y aminoácidos libres.
Las enzimas del ribete en cepillo tienen una distribución diferencial dentro de la vellosidad. Este hecho se asocia con diferencias en la estabilidad o labilidad de las mismas. Por ejemplo, la sacarasa se distribuye en toda la superficie vellositaria y es muy estable ante las agresiones. De allí que los factores que pueden dañar la mucosa difícilmente afecten la hidrólisis de la sacarosa. En cambio la lactasa se ubica en la porción distal de la vellosidad y su estructura es más lábil. Este hecho explica la facilidad con que la enzima disminuye y aun desaparece ante la acción de noxas (virus, bacterias, etc.).
En ciertas etnias, los individuos adultos dejan de expresar la lactasa y se hacen intolerantes a la lactosa. La ausencia de lactasa en el ribete en cepillo los incapacita para digerir la lactosa, por lo que este azúcar permanece en la luz intestinal ocasionando diarrea.
Después de su paso por el intestino delgado, todos los componentes del alimento han sido expuestos a la totalidad de las enzimas presentes, de manera que alcanzan el máximo grado de hidrólisis que dichas enzimas posibilitan, y están en condiciones de ser absorbidos.
Además de la digestión química, el intestino delgado realiza movimientos musculares que toman parte en la digestión mecánica. En el intestino delgado se producen movimientos de segmentación y movimientos de avance (peristálticos).
Los movimientos de segmentación ayudan a la mezcla del contenido luminal. Al mezclar el quimo con la bilis, el jugo pancreático y las secreciones intestinales, aumentan la interacción de los componentes del alimento con las sales biliares y las enzimas. También favorecen el contacto con la mucosa intestinal. Esto facilita la digestión de superficie, a nivel del ribete en cepillo, y la posterior absorción de los productos de la digestión.
%20de%20seg-3.JPG) |
Los movimientos de segmentación consisten en la contracción rítmica alternada de distintas partes del tubo y permiten la mezcla del contenido intestinal. |
Los movimientos peristálticos propulsan el contenido intestinal en sentido distal. Conforme ocurre el avance, los nutrientes en estado monomérico son absorbidos por las vellosidades intestinales.
Resumen de la digestión química a lo largo del tubo digestivo Regresar
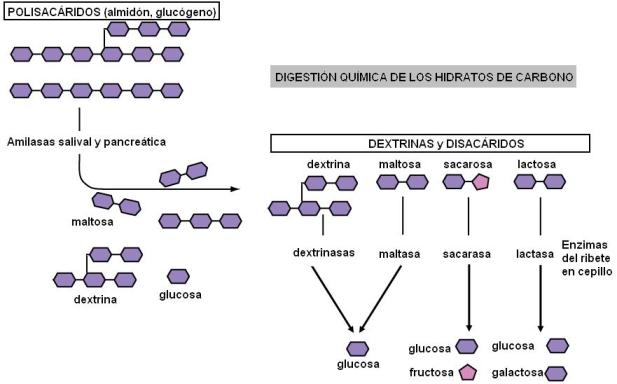
• Hidratos de carbono. La digestión química de los polisacáridos se inicia en la boca, por acción de la ptialina o amilasa salival. Como producto de la hidrólisis se obtienen maltosa, dextrinas, trisacáridos y en pequeña medida, glucosa libre. La acción de la amilasa salival se inhibe cuando el bolo alimenticio se mezcla con el jugo gástrico en la formación del quimo. En el estómago no hay hidrólisis de los hidratos de carbono. Los polisacáridos que no pudieron hidrolizarse en la boca son atacados en el duodeno por la amilasa pancreática, la cual posee la misma acción que la amilasa salival. Las dextrinasas y disacaridasas del ribete en cepillo completan la hidrólisis hasta la obtención de monosacáridos.
 |
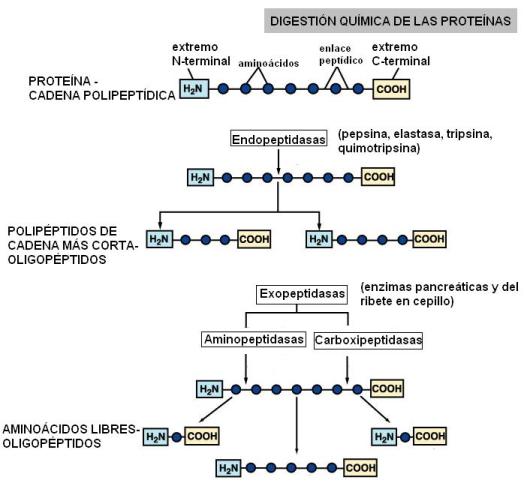
• Proteínas. La hidrólisis de las proteínas comienza en el estómago, por acción de la pepsina. Ésta escinde a las cadenas polipeptídicas, hidrolizando enlaces del interior de la cadena (acción endopeptidasa); genera polipéptidos de menor longitud u oligopéptidos. En duodeno continúa la digestión por acción de las endopeptidasas y carboxipeptidasas pancreáticas, las que liberan oligopéptidos y aminoácidos libres. Los oligopéptidos son atacados finalmente en la superficie del ribete en cepillo por las aminopeptidasas, que liberan aminoácidos del extremo aminoterminal y por las dipeptidasas. Los productos finales de la hidrólisis son tripéptidos, dipéptidos y aminoácidos libres.
 |
• Lípidos. Los lípidos no son atacados químicamente hasta que llegan al duodeno. Allí se emulsionan, por acción de las sales biliares. Una vez formadas las micelas, la colipasa pancreática ayuda al anclaje de la lipasa, que hidroliza los enlaces éster de los triglicéridos. Los productos de la hidrólisis permanecen en la micela hasta que hacen contacto con el ribete en cepillo para su absorción.
Absorción de los glúcidos Regresar
Los productos finales de la digestión de los glúcidos son los monosacáridos. El más abundante de ellos es la glucosa, pues es el monómero de los glúcidos más representados en la dieta: los almidones. Otros monosacáridos son la fructosa, proveniente del azúcar de caña y la galactosa, proveniente de la leche.
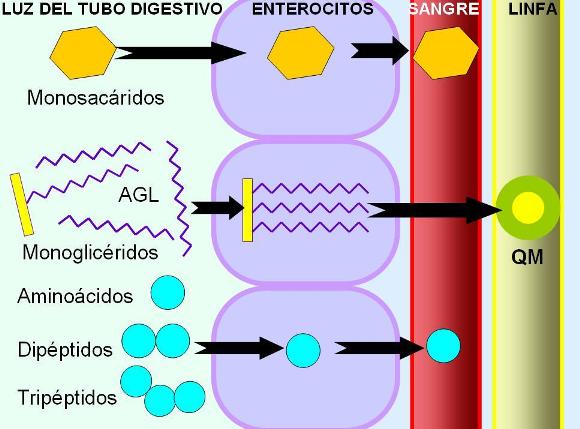
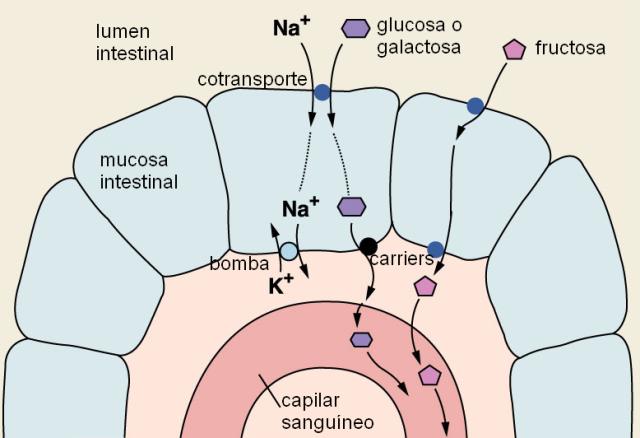
La absorción de la glucosa se realiza por un mecanismo de cotransporte con Na+. En la superficie apical de los enterocitos se encuentra un cotransportador Na+/glucosa. El cotransportador es una proteína con dos sitios de unión específicos: uno para el Na+ y otro para la glucosa. Cuando ambos solutos se unen al transportador en la cara luminal de la membrana, se produce un cambio conformacional del transportador, con la consecuente liberación del Na+ y la glucosa hacia el citosol. El Na+ ingresa a favor de su gradiente de concentración, en tanto la glucosa, más concentrada en el medio intracelular que en el extracelular, ingresa contra su propio gradiente, arrastrada por el Na+. Este mecanismo es considerado un transporte activo secundario, ya que si bien el transportador no requiere la hidrólisis de ATP en forma directa, depende de la existencia del gradiente de Na+. El gradiente de concentración es una forma de energía potencial acumulada que, en última instancia, se sostiene por el continuo accionar de la bomba de Na+ / K+, es decir, con consumo de ATP. La bomba de Na+/K+ está ubicada en la superficie basolateral de las células intestinales.
La existencia de un mecanismo de transporte activo garantiza que la glucosa, un nutriente fundamental, se incorpore al enterocito aun cuando su concentración luminal sea menor que la intracelular.
Una vez dentro del enterocito, la glucosa lo abandona rumbo a la circulación sanguínea a través de la superficie basal. En la superficie basal del enterocito se encuentra un carrier para glucosa que facilita su difusión hacia el espacio intersticial.
Las uniones estrechas u oclusivas en las superficies laterales de los enterocitos impiden el desplazamiento de los transportadores de la membrana apical hacia la basolateral y viceversa. Esta restricción es indispensable para mantener la polarización de la célula, garantizando así el transporte unidireccional de la glucosa desde la luz intestinal hasta la sangre. Téngase en cuenta que si el carrier para la difusión facilitada de glucosa llegara a la superficie apical, ésta se movería según su gradiente, retornando a la luz del intestino, con su consecuente pérdida.
La galactosa es transportada mediante cotransporte con Na+, por el mismo mecanismo que la glucosa. La fructosa, en cambio, es absorbida por difusión facilitada. Todos los monosacáridos son transportados por la vena porta.
 |
Absorción de los productos de la digestión proteica Regresar
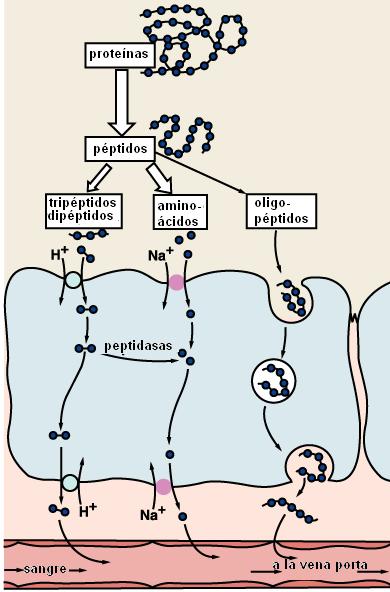
Como ya mencionáramos, las proteínas son degradadas hasta aminoácidos libres, dipéptidos y tripéptidos.
Los aminoácidos son absorbidos mediante cotransporte con sodio y por difusión facilitada.
Los tripéptidos y dipéptidos se absorben más rápidamente que los aminoácidos. Los pequeños péptidos ingresan al enterocito mediante un mecanismo de cotransporte con sodio. Dentro de la célula intestinal son digeridos a aminoácidos libres.
Los únicos oligopéptidos conocidos que se absorben son los provenientes de la gelatina (colágeno) y de ciertas carnes.
Normalmente no se absorben proteínas completas. Sin embargo, en ciertas circunstancias, cadenas polipeptídicas pueden ingresar mediante transcitosis (transporte en masa). Por ejemplo, en el recién nacido, éste es un mecanismo fisiológico para incorporar las inmunoglobulinas maternas secretadas con el calostro.
En las alergias alimentarias existe una reacción inmunológica contra proteínas presentes en los alimentos. El hecho de que exista esta sensibilización implica que las proteínas, o al menos fragmentos de la cadena polipeptídica, debieron alcanzar el medio interno sin previa hidrólisis.
Los productos de la digestión de las proteínas alcanzan el torrente circulatorio desde la superficie basal del enterocito, ingresando a la circulación portal.
 |
Absorción de los lípidos Regresar
La digestión y absorción de los lípidos depende de la formación de las micelas. Las vitaminas liposolubles se absorben con el resto de los lípidos. Cualquier trastorno que afecte la digestión y absorción de las grasas de la dieta provocará un déficit de estas vitaminas.
Los productos de la digestión de los lípidos y las vitaminas liposolubles se liberan de las micelas en contacto con la superficie apical del enterocito. Los ácidos grasos libres, los monoacilglicéridos, el glicerol, el colesterol y las vitaminas liposolubles atraviesan la membrana plasmática por difusión simple. Las sales biliares son recicladas.
Dentro del enterocito, los lípidos pasan al retículo endoplasmático liso (REL), donde gran parte de ellos se vuelven a esterificar, resintetizándose los triglicéridos, fosfolípidos y ésteres de colesterol. Desde el REL los lípidos son transportados hacia el aparato de Golgi. Combinados con la apoproteína beta, los lípidos forman los quilomicrones.
Los quilomicrones se exocitan por la superficie basal de las células intestinales y acceden a los capilares linfáticos o quilíferos.
Los ácidos grasos de cadena corta y media requieren menor trabajo digestivo que los de cadena larga y pueden ser absorbidos directamente hacia la circulación portal. Por este motivo son utilizados en alimentaciones especiales, cuando existen trastornos en la digestión y/o absorción de las grasas.
 |
Absorción de vitaminas hidrosolubles y minerales Regresar
Las vitaminas hidrosolubles son absorbidas en la parte alta del intestino delgado, excepto la B12, que se absorbe en el íleon. La absorción de la vitamina B12 requiere la unión de la vitamina con el factor intrínseco, secretado por las células parietales gástricas.
El calcio y el hierro también son absorbidos en la primera parte del intestino delgado. La absorción intestinal de calcio es facilitada por la vitamina D y se ajusta a las necesidades del organismo, aumentando cuando hay una disminución del Ca2+ corporal y disminuyendo cuando aquél se incrementa.
El hierro hémico (proveniente de la hemoglobina y la mioglobina) se absorbe más fácilmente que el no hémico. Este último, que generalmente se encuentra al estado férrico (Fe3+) en los alimentos debe reducirse al estado ferroso (Fe2+) para ser absorbido. La combinación de los alimentos que contienen hierro con vitamina C (ácido ascórbico), ayuda a la conversión del hierro férrico a ferroso, mejorando la absorción.
Absorción de agua y electrólitos Regresar
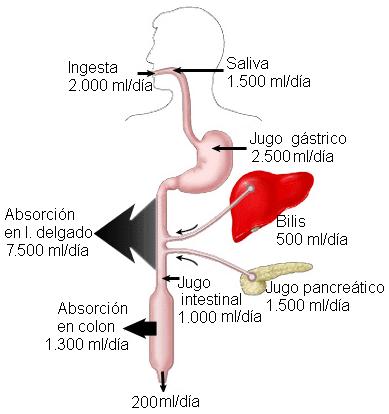 |
El intestino recibe diariamente unos 2 litros de líquido con la ingesta, más 7 litros provenientes de las secreciones de los órganos digestivos y las glándulas anexas. El 98 % de este líquido se reabsorbe, con una pérdida neta de 200 ml a través de las heces.
La mayor parte del líquido es reabsorbida en el intestino delgado.
El agua se mueve en ambas direcciones a través de la mucosa del intestino delgado en respuesta a gradientes osmóticos. El agua se desplaza hacia la luz intestinal o hacia el medio interno, hasta que la presión osmótica del contenido intestinal iguala a la del plasma. Dependiendo de la comida ingerida, la osmolaridad del contenido duodenal puede ser hipotónica o hipertónica; sin embargo, al llegar al yeyuno, la osmolaridad en la luz ya es igual a la del plasma.
El sodio es transportado activamente por la bomba de Na+/K+ en la membrana basolateral de los enterocitos. Esto mantiene un gradiente desde el lumen hacia el medio interno, necesario para propulsar el transporte de azúcares, péptidos, aminoácidos y otros nutrientes. De forma recíproca, la presencia de glucosa en el lumen favorece la absorción de sodio. Esta relación es la base fisiológica de la terapéutica de rehidratación oral utilizada en las diarreas, cuando se produce una pérdida de agua y sodio. Se suministra entonces una solución de sales de rehidratación oral, que contienen cloruro de sodio y glucosa. La absorción de sodio y glucosa se potencian y se absorbe agua por arrastre osmótico, restableciendo así el equilibrio hidroelectrolítico.
 |
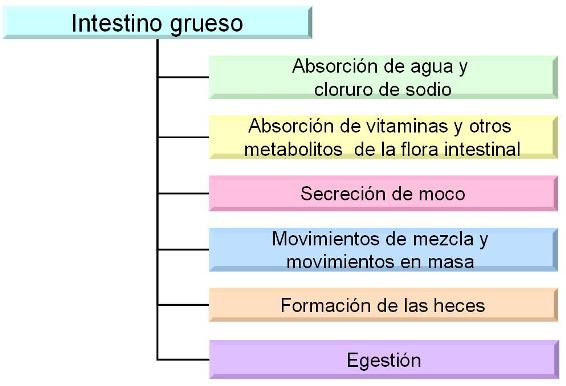
Anatomía y fisiología del intestino grueso Regresar
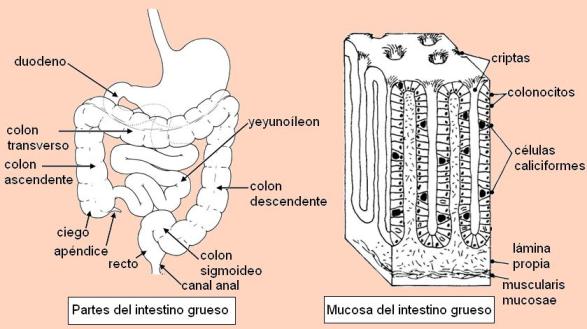
El intestino grueso se extiende desde la válvula ileocecal hasta el ano y tiene una longitud de 1,5 m. Consta del ciego, el apéndice, el colon, el recto y el canal anal.
El ciego está conectado con el apéndice, el cual tiene agregados de tejido linfático. El apéndice tiende a obstruirse e inflamarse, exigiendo su extirpación quirúrgica para evitar su rotura y una peritonitis.
El colon se divide en las siguientes porciones: colon ascendente, colon transverso, colon descendente y colon sigmoideo o ileopélvico; dichas porciones describen un marco alrededor de las asas intestinales.
El intestino grueso es de mayor calibre que el delgado, pero más corto que éste.
El músculo longitudinal del intestino grueso no forma una capa continua, sino que está dividido en bandas longitudinales, las tenias. Como dichas bandas son más cortas que la capa muscular circular, provocan la formación de unos pliegues (haustras) en la pared del intestino grueso.
El canal anal posee dos esfínteres. El interno es un engrosamiento del músculo liso circular y su control es involuntario. El esfínter externo, de control voluntario, está formado por músculo estriado.
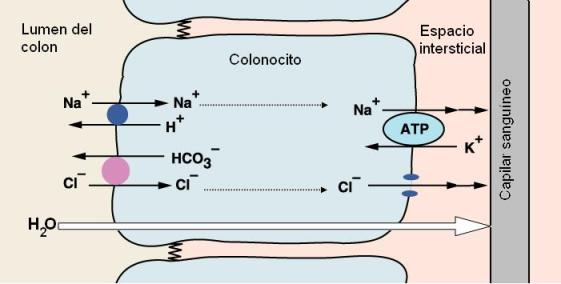
La mucosa del intestino grueso no presenta pliegues. Contiene numerosas glándulas tubulares rectas con células caliciformes. Las células de revestimiento, los colonocitos, están especializadas en la absorción de agua y electrólitos.
 |
Al colon no llegan nutrientes orgánicos, ya que éstos se absorben en el intestino delgado. El colon recibe alrededor de 1,5 l de líquido espeso, que se transforman en unos 250 cc de materia fecal semisólida, de manera que la mayor parte del agua que alcanza el colon es reabsorbida. Entre las materias sólidas se encuentran bacterias, células descamadas y fibra. La fibra incluye hidratos de carbono y otros polímeros indigeribles como celulosa, hemicelulosa y lignina.
Como el contenido del colon se va solidificando a medida que avanza, se secreta gran cantidad de moco, que lo protege de posibles traumatismos.
La reabsorción de agua se realiza por arrastre osmótico, siguiendo al sodio y al cloruro, que son absorbidos mediante mecanismos activos.
El sodio y el cloruro ingresan por la membrana apical del colonocito, mediante sendos antiportes. El sodio se intercambia por protones y el cloruro por bicarbonatos. Luego, sodio y cloruro son transportados hacia el líquido intersticial por la superficie basolateral. La acumulación de estos cationes en el líquido intersticial torna a éste hipertónico con respecto al lumen. En virtud del gradiente osmótico, el agua es arrastrada hacia el espacio intersticial en contacto con la superficie basolateral del epitelio. De esta forma sodio, cloruro y agua llegan por el espacio intersticial hacia los capilares sanguíneos.
 |
En el colon se producen contracciones que mezclan su contenido y permiten que el mismo haga contacto con la superficie mucosa, favoreciendo la reabsorción de agua y sales.
Los movimientos peristálticos, llamados movimientos en masa, se producen entre 1 y 3 veces por día y empujan el contenido del colon hacia el recto.
El reflejo gastroduodenocólico consiste en un aumento del peristaltismo del colon, como respuesta al llenado gástrico o a la recepción del quimo en el duodeno. El contenido fecal es barrido hacia el recto, preparando la defecación.
El volumen fecal estimula el peristaltismo, de allí la importancia del consumo de fibra para mantener una defecación regular.
La necesidad de defecar se percibe cuando el contenido fecal distiende las paredes del recto. Este estímulo relaja temporalmente el esfínter interno. Si el esfínter externo no se relaja voluntariamente, el esfínter interno se contrae nuevamente y la urgencia de la necesidad de defecar desaparece por un tiempo.
Diarrea Regresar
La diarrea se caracteriza por una deposición acuosa, es decir un aumento del contenido líquido de las heces.
La diarrea puede obedecer a una falla en la absorción de algún nutriente o electrólito osmóticamente activos. Los nutrientes no absorbidos atraen agua hacia el lumen, ocasionando la eliminación de mayor cantidad de agua en las heces (diarrea osmótica).
Este tipo de diarrea también ocurre cuando se produce un desarreglo alimentario y el flujo del contenido intestinal se ve aumentado, sin dar tiempo suficiente a la absorción.
Otras veces, la secreción en las criptas de Lieberkühn es estimulada por algún factor (acetilcolina, serotonina, toxina colérica) superando la capacidad de reabsorción de las vellosidades (diarrea secretora).
Flora del aparato digestivo Regresar
El aparato digestivo está colonizado por bacterias, en su mayoría comensales, que en conjunto forman la flora. Los únicos sitios relativamente estériles son las glándulas salivales, la vesícula biliar y el estómago. Cuando hay aclorhidria (el estómago no secreta ácido) existe una predisposición a las infecciones intestinales.
Helycobacter pylori está adaptada al medio ácido y se encuentra en el antro gástrico y en el duodeno; esta bacteria tiene un importante papel en la etiología de las úlceras pépticas.
En el intestino grueso hay al menos 400 especies de bacterias. Las bacterias forman alrededor de la cuarta parte del peso de las heces.
La flora del intestino proporciona vitaminas K y B12, que pueden ser absorbidas.
En el intestino grueso la flora transforma la bilirrubina en urobilinógeno (que en parte se reabsorbe y se elimina por orina) y estercobilinógeno, el cual se elimina con la materia fecal. El color pardo de las heces se debe a estos pigmentos.
El metabolismo bacteriano de la fibra también genera diversas sustancias absorbidas en el colon (amoníaco, ácidos grasos de cadena corta y aminas), otras responsables del olor de la materia fecal (como el escatol) y contribuye a formar los gases de los flatos.
 |
|
|