| INTRODUCCIÓN AL ESTUDIO DE LA CÉLULA Silvia Márquez - Lionel Valenzuela Pérez - Gladys Gálvez - Luis A. Fernández - Cecilia Bocchino NIVELES DE ORGANIZACIÓN
A los seres vivos se los define por sus características, una de éstas es su organización. Esta organización biológica representa el patrón complejo que nos muestra el camino que ha seguido la evolución, desde formas sencillas a otras más complejas. Los seres vivos están formados por materia. La materia está formada por elementos (92 elementos naturales, como el Cloro, por ejemplo) y se caracteriza por poseer determinadas propiedades intensivas, tales como el punto de fusión, punto de ebullición, conductividad eléctrica, etc. Los elementos están formados por átomos. Un átomo es la porción más pequeña de un elemento que conserva sus propiedades químicas. Las investigaciones de los físicos han descubierto un variado número de partículas subatómicas (Nivel Subatómico), para nuestros fines mencionaremos sólo tres : protones, neutrones y electrones. Los protones son partículas con carga positiva; los electrones, en cambio, tienen carga negativa y masa muy pequeña; los neutrones son partículas neutras, sin carga, y su masa es casi idéntica a la de los protones; los protones y neutrones forman casi toda la masa de un átomo y se localizan en el núcleo atómico. Si combinamos un protón y un electrón se forma un átomo de Hidrógeno, entidad con propiedades diferentes a las de un protón y un electrón (Nivel Atómico). Si combinamos átomos de Hidrógeno entre sí obtenemos Hidrógeno molecular (H2), que es un gas incoloro; si, en cambio, combinamos el H2 con Oxígeno, otro gas, obtenemos agua, una molécula (Nivel Molecular) cuyas propiedades todos conocemos y que no son las mismas que las del H2 y el O2 y que también difieren de las propiedades de las partículas subatómicas y de los átomos que éstas forman. La vida surgió a partir de átomos y moléculas. Si combinamos moléculas entre sí, formamos grandes y complejas moléculas: las macromoléculas, como las proteínas y los ácidos nucleicos (Nivel Macromolecular). Estas macromoléculas constituyen la materia prima que forman los virus (Nivel Prebiótico o Supramolecular) y las células (Nivel Celular). En el Nivel Subcelular múltiples moléculas se ensamblan y dan lugar a estructuras especializadas como los organoides (mitocondrias, cloroplastos, etc). Podemos decir que la vida aparece como propiedad definitoria en el Nivel Celular, o de otro modo, la célula es la porción más sencilla de la materia viva que es capaz de realizar todas las funciones imprescindibles para la vida. En la mayor parte de los individuos pluricelulares, las células se organizan de acuerdo a sus características y funciones conformando tejidos como el conectivo, muscular, epitelial, nervioso (Nivel Tisular). Los tejidos están ordenados en estructuras funcionales, denominadas órganos como el corazón y los pulmones en los animales, o las hojas y las raíces en las plantas. Las funciones biológicas básicas se llevan a cabo por un sistema o aparato, que es una asociación coordinada de tejidos y órganos. Los organismos o individuos pluricelulares están formados por sistemas que actúan en forma coordinada y rigurosa. Existen otros niveles de organización biológica, además de los nombrados anteriormente, donde las propiedades provienen de la relación entre los organismos. Por ejemplo, el Nivel de organización POBLACIÓN reúne a todos los individuos de una misma especie que viven en un mismo lugar, en el mismo tiempo, y que comparten el mismo hábitat. Estas poblaciones interactúan de distinta manera con otras poblaciones del lugar constituyendo una COMUNIDAD, por ejemplo la población de seres humanos de la ciudad de Buenos Aires y el conurbano, aprovecha para alimentarse a las distintas poblaciones de animales y plantas de la zona y se halla parasitada por las mismas poblaciones de parásitos intestinales. Esta comunidad comparte el mismo lugar físico que presenta características particulares. La unión de estos factores físicos con los factores biológicos constituyen los ECOSISTEMAS. Todos los ecosistemas de la Tierra están relacionados, directa o indirectamente. Es por ello que un cambio drástico o continuo de alguno de ellos indefectiblemente acarreará cambios en los restantes. Del mantenimiento de un equilibrio entre los distintos ecosistemas, depende la vida en el planeta.
Durante el desarrollo de nuestra materia, nos ocuparemos de los niveles de organización más sencilla y haremos hincapié en el Nivel Celular de Organización.
CaracterÍsticas de los seres vivos En la mayoría de los textos de Biología no hay una definición de la vida sino de los seres vivos, es decir que a través de sus características o propiedades podemos distinguir la materia inerte de la materia viva. ClASIFICACIÓN DE LOS SERES VIVOS
La clasificación de los seres vivos se realiza según relaciones evolutivas (Tabla 1.3). Este tipo de clasificación se denomina sistemática.
OrganizaciÓn Celular Teoría Celular La célula es la unidad de vida más pequeña. Es la unidad anatómica y fisiológica de todos los seres vivos. Dos científicos alemanes el botánico Mattias Schleiden (1804-1881) y el zoólogo Theodor Schwann (1810-1882) fueron los primeros en señalar que "Los cuerpos de las plantas y de los animales están compuestos por células y por productos celulares" enunciando el postulado inicial de la Teoría Celular Posteriormente, Rudolph Virchow (1821-1902) amplio la Teoría Celular y afirmó: "Todas las células proceden de otra preexistente". Por lo tanto, las células no surgen por generación espontánea a partir de materia inanimada. Otra importante conclusión de la Teoría Celular afirma que todas las células actuales, tienen un origen común. La evidencia más importante, sobre el origen común de todas las formas celulares, radica en las similitudes básicas de sus estructuras y principalmente de su composición molecular.
CARACTERÍSTICAS DE LAS CÉLULAS Todas las células están cubiertas por una membrana externa, llamada membrana plasmática, que las separa de otras células y del medio circundante con el cual intercambian materia y energía. Este intercambio esta altamente regulado y es selectivo. De esta forma la membrana plasmática debe actuar no sólo como limite celular sino también como barrera selectiva. Por lo tanto la célula, mantiene una composición química muy ordenada y diferente a la del entorno. Todas las células poseen un metabolismo o conjunto de reacciones químicas, que posibilitan el mantenimiento de la vida. Este metabolismo para sustentarse necesita de una o más fuentes de energía. Las células, necesitan de distintivos tipos de moléculas energéticas: * Monedas energéticas, como el ATP * Moléculas combustibles, como la glucosa o los ácidos grasos * Moléculas de reserva de energía, como el glucógeno o el almidón Dentro de las reacciones para obtener e interconvertir diferentes forma de energía, son muy importantes las reacciones de oxido-reducción o reacciones REDOX. En este tipo de reacciones es esencial la participación de las coenzimas de oxido-reducción, como el NAD+ y el FAD. Todas las células, almacenan en forma de ADN, ácido desoxirribonucleico, a información necesaria para controlar sus actividades (reproducción, metabolismo), y para establecer su propia estructura. El ADN, es un polímero formado por una secuencia lineal, de monómeros, llamados nucleótidos. Esta secuencia de nucleótidos, especifica una secuencia de aminoácidos (estructura primaria de una proteína). La especificidad de la secuencia de aminoácidos determinada por la secuencia de bases del ADN esta regida por el código genético. La secuencia de bases del ADN, que codifica una proteína, es un GEN. Las proteínas, son moléculas que llevan a cabo gran parte de las funciones celulares. Muchas proteínas son enzimas, moléculas encargadas de dirigir y regular el metabolismo celular. Las enzimas aceleran las reacciones químicas, haciéndolas compatibles con la vida. De esta manera las enzimas, dirigen la síntesis y degradación de todas las moléculas biológicas, incluidos lípidos, glúcidos, proteínas y los mismos ácidos nucleicos. De esta forma, el ADN al almacenar la estructura de las enzimas y otras proteínas reguladoras, ejerce el control del metabolismo celular. El ADN utiliza un segundo ácido nucleico, el ARN, ácido ribonucleico, como intermediario. A partir de la secuencia de bases del ADN, que codifica una proteína, se sintetiza una secuencia de bases de ARN. Este proceso es llamado transcripción. EL ácido ribonucleico encargado de transportar la información, recibe la denominación de ARN mensajero. Este ARN mensajero, porta la información necesaria para la síntesis de proteínas, proceso llamado traducción, el cual tiene lugar en el citoplasma con la intervención de dicho ARNm, los ribosomas y el ARNt que porta los aminoácidos. Las células para perpetuarse necesitan reproducirse. Esto significa que la información almacenada en el ADN debe duplicarse para poder ser transmitida a las células hijas. El ADN tiene la excepcional característica de ser una molécula capaz de autorreplicarse, es decir de generar una copia de si misma. Este proceso es llamado duplicación o replicación. DIMENSIONES DE LAS CÉLULAS ¿Por qué son tan pequeñas las células? Las células deben captar alimento y otros materiales a través de su membrana plasmática y deben eliminar los productos de desecho, generados en las distintas reacciones metabólicas rápidamente antes de que estos se acumulen hasta niveles tóxicos para la supervivencia celular. Por lo tanto, las células son pequeñas, de modo que en ellas las moléculas recorren distancias cortas, lo que acelera las actividades celulares. Además, a mayor superficie celular, mayor es el transporte de moléculas a través de la membrana, siendo importante para la continuidad de los procesos metabólicos la proporción superficie celular sobre volumen celular. Supongamos una célula de forma cúbica, cuanto más grande es, su superficie crece proporcionalmente lado x lado, es decir a la segunda potencia de la longitud de un lado, en cambio el volumen celular aumenta proporcionalmente a la tercera potencia. Por lo tanto, el volumen celular aumenta más que su superficie a medida que la célula crece, determinando el limite superior al tamaño de la célula en cuestión. Está célula sólo podrá iniciar el proceso de división celular (previa duplicación de su ADN) o perecerá. Por otra parte, debemos recordar que en las células el material Genético (localizado en el núcleo, en células eucariontes), posee un área limitada de influencia sobre el citoplasma circundante, que es el que incrementa marcadamente su tamaño durante el crecimiento celular, siendo otra limitante del tamaño celular la relación núcleo/citoplasma. CÉLULAS EUCARIOTAS Y PROCARIOTAS CARACTERÍSTICAS PRINCIPALES Todas las células se parecen y responden a un patrón común por más diversas que sean. Las células de organismos pluricelulares son diferentes en su función, por ser distintas estructuralmente, pero todas concuerdan con un patrón común. Por ejemplo, aquellas especializadas en la síntesis de lípidos, tendrán mayor desarrollo del retículo endoplasmático liso y serán distintas de las neuronas especializadas en la transmisión del impulso nervioso, cuya especialización es tan grande que pierden su capacidad de reproducirse. A pesar de las semejanzas y diferencias entre las células y que todas cumplen con los postulados de la Teoría Celular, se distinguen dos grandes tipos de células: PROCARIOTAS (sin núcleo verdadero) y EUCARIOTAS (con núcleo).
Los eucariontes son organismos cuyas células poseen un sistema de endomembranas (membranas internas) muy desarrollado. Estas membranas internas forman y delimitan organelos donde se llevan a cabo numerosos procesos celulares. De hecho él más sobresaliente de estos organelos es el núcleo, donde se localiza el ADN. Justamente, el término eucarionte, significa núcleo verdadero (eu: verdadero, carion: núcleo). Por lo tanto, las células eucariontes, poseen diversos compartimentos internos, rodeados por membranas. De esta forma es más eficiente reunir a los sustratos y sus enzimas, en una pequeña parte del volumen celular total. Además de conseguirse una mayor velocidad, las membranas favorecen la aparición de estructuras reguladoras que orientan el flujo de moléculas y su posterior conversión en otros productos. Ciertos procesos como la fotosíntesis y la cadena respiratoria están altamente organizados gracias a la localización de las enzimas en diferentes estructuras de membrana. Por otra parte, las membranas también impiden la aparición de sustratos en forma inespecífica en distintas regiones de la célula, ya que actúan como barrera selectiva. En cuanto al tamaño, podemos decir que en promedio una célula eucarionte es diez veces mayor que una célula procarionte. En cuanto al material genético, podemos decir que el ADN eucariota posee una organización mucho más compleja que el ADN procarionte. Las células procariontes carecen de núcleo y generalmente son mucho menores que las células eucariontes. El ADN de las células procariontes no está rodeado por una membrana, pero puede estar limitado a determinadas regiones denominadas nucleoides. Las células procariontes, al igual que las células eucariontes, poseen una membrana plasmática, pero carecen de membranas internas, que formen organelos. Sin embargo, debemos precisar que en algunas células procariontes, la membrana plasmática forma laminillas fotosintéticas. Las células procariontes poseen una característica única, una pared de peptidoglicanos, un gran polímero de glúcidos y aminoácidos.
Estructura de las células PROCARIóTicAS Fig. 1.2- Esquema de una ultraescructura de una bacteria idealizada Bacterias, Micoplasmas y Algas Cianofíceas Cuadro 1.1- Estructura de una Célula procarióta Las bacterias pueden definirse como organismos unicelulares procariontes que se reproducen por fisión binaria. Contienen toda su información genética en un único cromosoma bacteriano circular. También poseen sistemas productores de energía y biosintéticos necesarios para el crecimiento y la reproducción. Poseen como característica particular una pared rígida de peptidoglicanos. Son generalmente de vida libre y poseen ADN extracromosómico en forma de plásmidos, estos codifican genes de resistencia a antibióticos o factores "sexuales" como los pili. Los micoplasmas son las bacterias mas pequeñas de vida independiente. Son muy flexibles y deformables por lo que atraviesan los filtros de esterilización. Entre sus características principales se encuentran: a) carecen de pared celular, b) en su membrana plasmática poseen esteroles, que no son sintetizados por la bacteria sino que son absorbidos del medio de cultivo o del tejido donde se desarrolla.. Los micoplasmas son resistentes a la penicilina (carecen de pared de peptidoglucano) y por la misma razón no toman la coloración de Gram. Las cianobacterias, anteriormente llamadas algas cianofíceas (azulverdosas), son bacterias Gramnegativas. Se encuentran presentes en estanques, lagos, suelo húmedo, cortezas de árboles, océanos y algunas en fuentes termales. La mayor parte de las cianobacterias son autótrofos fotosintéticos. Contienen clorofila a, que también se encuentra en plantas y algas. La clorofila a y pigmentos accesorios se localizan en membranas fotosintéticas, llamadas laminas internas o laminillas fotosintéticas. Muchas especies de cianobacterias fijan nitrógeno, este proceso enriquece el suelo. En la Tabla 1.4 se aprecian las diferencias más importantes entre las células procariotas (bacterias) y eucariotas Plásmidos Un plásmido es una molécula de ADN extracromosómico que se replica en forma autónoma, por lo que al igual que el cromosoma es un replicón. Puede haber hasta 50 copias de un plásmido en una bacteria. Funcionalmente los plásmidos son elementos genéticos accesorios, es decir que la bacteria puede vivir sin ellos. Sin embargo, la información que contienen puede contribuir a la adaptación de la bacteria al medio y a la evolución de la misma. Los plásmidos pueden contener genes que codifican factores de resistencia a antibióticos (los plásmidos R) y factores de patogenicidad como exotoxinas. La evolución bacteriana a través de los plásmidos es factible, ya que pueden ser intercambiados entre distintas bacterias (por ejemplo, el plásmido F). Es decir que ciertos genes pueden transferirse de una bacteria otra mediante el pasaje de plásmidos, a este mecanismo se lo denomina conjugación. Para que la conjugación pueda llevarse a cabo las dos bacterias deben ponerse en contacto físico . Esto es posible debido a que una de las bacterias, posee pili sexuales (pelos) en su envoltura. Estos pili se encuentran codificados en el mismo plásmido F (plásmido conjugativo). La bacteria que transfiere el plásmido es la que posee pili y se la denomina F+, la célula receptora es F-. Transposones Los transposones son elementos genéticos movibles, que se encuentran presentes en los procariontes (aunque también en las células eucariontes). El descubrimiento de los transposones se lo debemos a Barbara McClintock. Los transposones son fragmentos de ADN que se mueven de una localización a otra del cromosoma. Esta transposición es catalizada por una enzima llamada transposasa. El gen de la transposasa esta incluido dentro del mismo transposón. Los transposones al ser elementos móviles, dentro del genoma, pueden provocar mutaciones al insertarse en nuevas regiones del ADN. Pared Celular. Bacterias Grampositivas y Gramnegativas Por fuera de la membrana celular, se encuentra una pared celular rígida de peptidoglicano, que esta presente en todas las bacterias excepto los micoplasmas. La presencia de la pared protege a la bacteria de la diferencia de presión osmótica entre el medio interno de la bacteria y el medio exterior. De no existir la pared la bacteria estallaría. Además la pared cumple funciones de protección como por ejemplo contra sustancias tóxicas . Existen dos tipos de pared bacteriana que pueden diferenciarse por la Tinción de Gram (siglo XIX). El primer grupo de bacterias son aquellas capaces de retener el colorante cristal violeta luego de la decoloración con alcohol-cetona. Estas bacterias son llamadas Grampositivas. El segundo grupo esta conformado por aquellas bacterias incapaces de retener el colorante luego del tratamiento decolorante, por lo tanto son llamadas Gramnegativas. La pared celular de las bacterias Grampositivas
La pared celular de las Grampositivas es más gruesa que la de los Gramnegativas. Posee peptidoglicano, ácidos teicoicos y lipoteicoicos. El componente fundamental es la mureína, un peptidoglicano que solo se encuentra en los procariontes. La mureína consiste en una cadena lineal de dos azúcares alternados N-acetilglucosamina y ácido acetilmurámico. A cada residuo de ácido murámico se encuentra unido un tetrapéptido compuesto de D- y L- aminoácidos. Aproximadamente un tercio de los tetrapéptidos presentes participan de la unión lateral entre cadenas adyacentes de mureína. La pared celular es biológicamente estable, resiste el ataque de las enzimas de los mamíferos, excepto de la lisozima que la degrada. La síntesis de la pared puede ser afectada por antibióticos como la penicilina. Los ácidos teicoicos son el principal determinante antigénico de las bacterias Grampositivas y por lo tanto definen la individualidad inmunológica de estas bacterias. La pared celular de las bacterias Gramnegativas
El espesor de la pared celular de una bacteria Gramnegativas es considerablemente menor que el de una Grampositivas. La cantidad de mureína es mucho menor en los Gramnegativas. Los ácidos teicoicos no están presentes en las bacterias Gramnegativas. A ambos lados de la fina pared de mureína se encuentra un gel periplásmico, que define al llamado periplasma (antes llamado espacio periplasmático). Por fuera del periplasma se encuentra una estructura exclusiva de las Gramnegativas, la denominada membrana externa. Si bien es estructuralmente similar a una bicapa lipídica, su composición es diferente de la de otras membranas biológicas. Esta bicapa es muy asimétrica, la semicapa interna esta compuesta por fosfolípidos, pero la semicapa externa esta compuesta por lipopolisacáridos (LPS), altamente tóxico para el ser humano (endotoxina). Para obtener nutrientes las bacterias Gramnegativas, poseen porinas que son proteínas que forman poros en la membrana externa. CÁpsulas Por fuera de la membrana externa de las Gramnegativas y de la gruesa pared de las Grampositivas, se encuentra presente, en algunas bacterias, una cápsula o matriz exopolisacárica, formada por un gel hidrofílico. En general esta cápsula o matriz esta formada por polímeros de azúcares. Las cápsulas permiten a las bacterias evadir los mecanismos de defensa de los organismos pluricelulares, también tienen funciones de adherencia a epitelios permitiendo de esta manera colonizar los tejidos del huésped. Estructura de las cÉlulas eucariÓticas
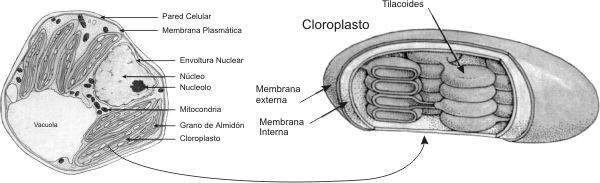
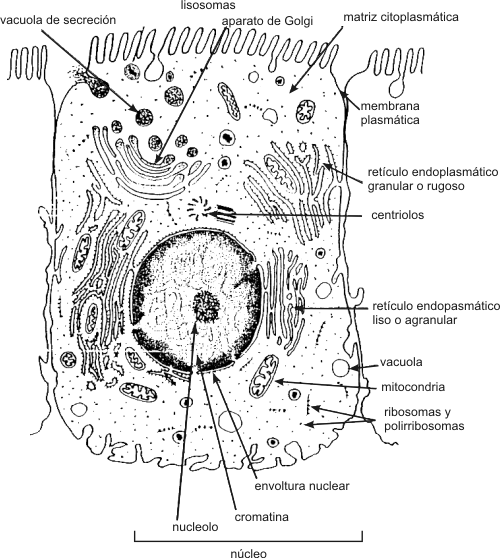
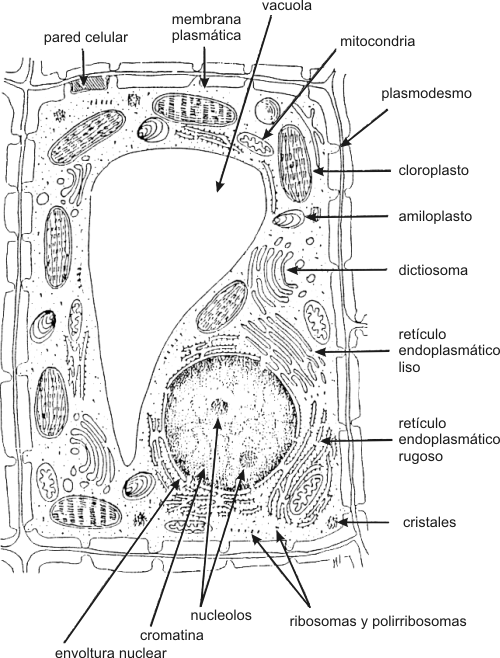
NÚcleo celular Las diversas partes de una célula eucariótica interactúan de forma integrada. Esto es posible porque existe un centro primordial de control: el núcleo celular. Una membrana doble, la envoltura nuclear (constituida por dos unidades de membrana), controla el transporte, muy selectivo, de sustancias entre el núcleo y el citoplasma. El pasaje se realiza a través de los poros nucleares. La envoltura nuclear posee ribosomas adheridos a la cara citoplasmática y una estructura proteica en su parte interna llamada lamina nuclear, que sirve como esqueleto al núcleo. En el interior del núcleo, se encuentra el material genético (ADN) asociado a proteínas básicas llamadas histonas, formando una estructura fibrilar muy enrollada denominada cromatina y el nucleolo, sitio de ensamblaje de los ribosomas (estructuras esenciales para la síntesis de proteínas, formados por ARN ribosomal y proteína). El ARN ribosómico se sintetiza en el nucleolo, y las proteínas ribosómicas en el citoplasma, para pasar después al núcleo y de allí al nucleolo, donde se unen al ARN ribosomal para formar los ribosomas.
Rodeando al núcleo encontramos el citoplasma, coloide donde predominan como constituyentes agua, iones, enzimas y donde se encuentran incluidos los organelos celulares. El citoplasma se encuentra separado del ambiente exterior por la membrana plasmática. Membrana plasmática Estructuralmente esta compuesta por una bicapa fosfolipídica. El colesterol esta presente en las células animales, pero esta ausente, en general, en plantas, hongos y procariontes (salvo micoplasmas). La membrana plasmática también contiene múltiples proteínas con diversas funciones. Podemos dividirlas en dos grandes grupos: a) proteínas integrales de membrana y b) proteínas periféricas de membrana. Las primeras atraviesan la membrana de lado a lado, mientras que las segundas están en contacto con la membrana, pero no la atraviesan. Algunas son enzimas reguladoras, otras receptores hormonales. Existen también proteínas transportadoras y canales reguladoras del movimiento de iones y moléculas a través de la membrana plasmática, de allí su enorme especificidad. Otra función importante de la membrana es la comunicación intercelular y el reconocimiento de diversos tipos de molécula (hormonas, virus, anticuerpos, toxinas, etc.) que interactúan con ella. En general esta función es llevada acabo por glucoproteínas y glucolípidos, que se encuentran solo en el lado externo de la membrana plasmática. Se cree que los glúcidos juegan un importante papel en la adhesión entre células. A esta capa, de glucolípidos y glucoproteínas se la denomina glucocálix. Sistema de endomembranas Este sistema se compone de sistemas membranosos interconectados entre sí, como el retículo endoplalmático liso o agranular (REL), el retículo endoplasmático rugoso o granular (REG) y el aparato de Golgi. Estas estructuras permiten la circulación de sustancias siempre dentro de formaciones limitadas por membrana interactuando por medio de vesículas.
Organelas
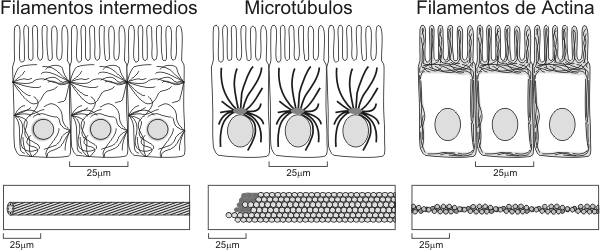
Ribosomas y Polirribosomas Son estructuras redondeadas que a diferencia de las anteriores, carecen de unidad de membrana. Están constituidos por dos subunidades, mayor y menor separadas entre sí. Ambas subunidades se unen cuando leen una molécula de ARNm. Las subunidades están formadas por ARNr y proteínas, siendo ensambladas en el nucleolo. Cuando hay varios ribosomas unidos a una molécula de ARNm, lo denominamos polirribosoma. La función de los ribosomas es sintetizar proteínas. Citoesqueleto El citoesqueleto es una red de fibras proteínicas. Esta red es dinámica encontrándose en constante cambio. Sus funciones, son esenciales para las células eucariontes y abarcan motilidad celular, forma, diferenciación, reproducción, regulación, etc.
CÉlula eucariÓta animal y vegetal
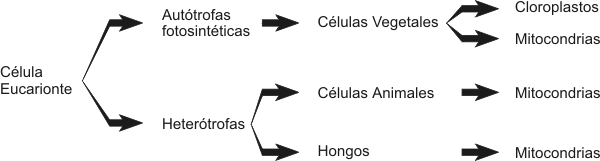
Las células eucariontes poseen dos modelos estructurales básicos: a) células autótrofas fotosintéticas y b) células heterótrofas. Las células autótrofas son aquellas que sintetizan su propio alimento, es decir sus propias moléculas combustibles. En este caso las células eucariontes vegetales son células autótrofas fotosintéticas, por lo tanto utilizan la luz solar como fuente de energía. Transforman la energía solar en energía química, este proceso es llamado fotosíntesis. La fotosíntesis en las células vegetales se lleva a cabo en un organelo membranoso llamado cloroplasto. Dentro del cloroplasto se encuentran sacos membranosos apilados, denominados tilacoides, en cuyas membranas encontramos el pigmento llamado clorofila, esencial para la fotosíntesis. Las células heterótrofas son aquellas que no sintetizan su propio alimento sino que necesitan una fuente externa de energía tanto como de materiales de construcción de sus propias moléculas. Las células animales (y los hongos), son células eucariontes heterótrofas. Las células animales y las células vegetales poseen unas organelas membranosas llamadas mitocondrias, donde se lleva acabo la respiración celular. En este proceso son rotos los enlaces de alta energía de las moléculas combustibles orgánicas. Esta energía liberada es utilizada para la síntesis de las monedas energéticas como el ATP. El ATP es esencial para las diferentes funciones celulares. Para que este proceso se lleve a cabo dentro de las mitocondrias es necesaria la presencia de oxigeno. Por lo tanto en ambos tipos celulares son necesarias las mitocondrias , para obtener energía química en forma de ATP a partir de las moléculas combustibles. Pero es diferente el origen de las moléculas orgánicas utilizadas como combustibles. En el caso de las células vegetales (autótrofas), ellas sintetizan sus propias moléculas combustibles en los cloroplastos, en el proceso de fotosíntesis. En cambio las células animales (heterótrofas), necesitan una fuente externa de moléculas energéticas que sirvan como combustible celular.
Fig. 1.8- Esquema de la ultraestructura de una célula
animal idealizada Fig. 1.9- Esquema de la ultraestructura de una célula vegetal idealizada
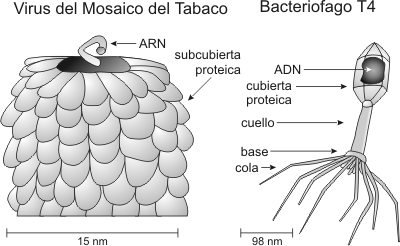
Hacia fines del siglo pasado se formulo la teoría de que cada enfermedad era producida por un germen específico. Hasta ese momento los patólogos estaban convencidos de que para cada enfermedad seria posible encontrar el microorganismo responsable, utilizando las siguientes técnicas: a) observación del germen con la ayuda del microscopio, b) cultivo sobre un medio nutritivo y c) retención por filtros. Sin embargo, en 1892, Iwanowski (o Ivanovsky?) pudo demostrar que el agente productor de la enfermedad del mosaico de tabaco pasaba a través de los filtros para bacterias y no podía ni verse ni cultivarse. Luego en 1898 Beijerinck, determinó que la enfermedad del mosaico del tabaco era provocada por un nuevo agente infeccioso a los que denomino virus filtrables (virus: palabra de origen latino que significa veneno). Los virus están ampliamente distribuidos en la naturaleza y afectan a todo tipo de organismos, tanto del reino animal, vegetal o protista. CaracterÍsticas estructurales de los virus La estructura de los virus esta integrada por dos tipos de macromoléculas: ácidos nucleicos y proteínas, los que forman las partículas virales o viriones. Básicamente existen dos tipos de partículas virales: partículas virales simples (virus desnudo) o partículas virales envueltas (virus envuelto). El virus desnudo consta de un ácido nucleico (genoma) asociado a proteínas y una cubierta proteica o cápside. Por otra parte los virus envueltos añaden a esta estructura básica una envoltura lipoproteica de origen celular. La función de la cápside es de servir al ácido nucleico como protección y vehículo. Postulado de Lwoff "Únicamente serán considerados virus aquellos agentes infecciosos cuya partícula elemental contenga un solo tipo de ácido nucleico". Es decir poseen ARN o ADN, pero no ambos tipos de ácidos nucleicos funcionales a la vez. Por lo tanto los virus pueden ser ADN o ARN virus. Debido a la estructura simple de virus, para su multiplicación dependen en forma absoluta de la célula huésped que infectan. Por lo tanto consideraremos a los virus como parásitos intracelulares obligados. ReplicaciÓn viral La célula huésped, una vez infectada, sintetizará nuevas moléculas de ácido nucleico viral, ya que el genoma viral tomara el control de las actividades metabólicas de la célula. Bajo este control, también se producirán gran cantidad de proteínas virales. De esta manera el ensamblado de nuevas partículas virales provendrá de la asociación de las nuevas moléculas de ácido nucleico viral con las proteínas. Este proceso es muy diferente de la reproducción celular, tanto de los procariontes como de los eucariontes. Por lo tanto es mas apropiado hablar de REPLICACION VIRAL. Los virus como parÁsitos intracelulares Hasta el momento no se ha podido demostrar que ningún virus aislado pueda utilizar o almacenar energía mediante procesos similares a la respiración, tampoco pueden sintetizar proteínas. Es decir no tienen metabolismo propio, dependiendo en forma absoluta del medio ambiente celular. El parasitismo de los virus se ejerce a nivel genético, ya que el genoma viral dentro de la célula desplaza al genoma de la célula hospedadora del control celular. Provirus Anteriormente hemos explicado que los virus, infectan las células y hacen replicas de si mismos. Luego abandonan la célula huésped y pasan a otra, recomenzando su ciclo. Pero también puede ocurrir que el genoma viral se integre al ADN del huésped. Cuando el genoma viral se integra al genoma celular y se replica junto con este se lo denomina PROVIRUS. Un provirus puede activarse espontáneamente o bien expuesto a diversos estímulos, una vez activado puede inducir la producción de virus completos. Los provirus pueden modificar la morfología celular y su metabolismo, esto puede deberse a la producción de alguna proteína viral. Estos cambios en la estructura celular, generalmente asociados a cambios en la membrana celular, inducidos por un provirus reciben el nombre de transformación. En algunos casos estas células transformadas por los provirus pueden ser cancerosas. Los virus como agentes infecciosos El parasitismo celular obligado es la causa básica por la cual un virus puede causar daño. La relación que establece un virus y su célula huésped es variable. El resultado de la interacción depende tanto del virus en cuestión como de la célula huésped. Por ejemplo, se denominan virus citocídicos , a los que como resultado de su multiplicación, producen una rápida inducción hacia la muerte celular. Por otra parte se encuentran los virus no citocídicos, que son aquellos que no provocan la muerte celular. Ejemplos de virus no citocídicos serian los virus moderados y los virus oncogénicos. Los virus moderados son aquellos que producen partículas virales y no producen la muerte celular. Han llegado con la célula huésped a un estado de equilibrio más o menos estable. Luego están los virus oncogénicos, capaces de estimular la división celular, estos cambios pueden ser irreversibles si la célula pierde la capacidad de regular su ciclo celular. Este estado se denomina transformación celular. Los virus no citocídicos pueden causar dos tipos de infecciones: 1- Las infecciones latentes, donde el virus permanece sin manifestarse, alojado en células no productoras. Ante determinados estímulos, estrés, enfermedades, exposición a la luz solar, el virus se reactiva, recomenzando la síntesis de ácidos nucleicos y proteínas virales. Ejemplos: L Herpes simplex, Varicela zoster. 2- Las infecciones crónicas, donde de una persona enferma siempre es posible obtener virus infeccioso, aun por periodos muy prolongados de tiempo. Por ejemplo, virus de la hepatitis B, Epstein-Barr, virus de la rubéola.
MorfologÍa viral Los virus poseen gran variedad de formas y tamaños. Por ejemplo los virus que al microscopio electrónico aparecen aproximadamente esféricos se denominan isométricos. En estos virus el ácido nucleico esta rodeado por una cápside (caja) proteica. Las subunidades estructurales que forman la cápside, visibles al microscopio electrónico, se denominan capsómeros. A su vez los capsómeros están formados por subunidades proteicas. La cápside por su naturaleza antigénica es la que determina la identidad viral. El ácido nucleico viral no se encuentra desnudo, sino que esta asociado a proteínas distintas de las de la cápside, cuya función puede ser estructural (plegado del ADN) o enzimática (polimerasa). Al conjunto de ácido nucleico y proteínas asociadas se lo denomina "core". Por ultimo diremos que los virus donde la cápside rodea directamente al ácido nucleico (es decir que no hay un "core" evidente), el conjunto de cápside y ácido nucleico recibe la denominación de nucleocápside. Genoma viral El genoma, que puede encontrase en los distintos tipos de virus, puede sistematizarse de acuerdo a diversos criterios: 1- Tipo de ácido nucleico: ADN o ARN 2- Polaridad o sentido: + o - (aplicado principalmente a los ARN virus) 3- Número de cadenas: monocatenario o bicatenario 4- Genoma circular o desnudo 5- Genoma entero o fragmentado Debemos recordar que las cadenas de un ácido nucleico de doble cadena, son de polaridad (sentido) opuesto. Por convención se considera que si una tiene sentido + la otra será -. De acuerdo a este criterio se considera que un virus es + (o cadena +), cuando su genoma monocatenario tiene la misma polaridad que un ARNm. Es decir el mismo genoma viral, puede actuar dentro de la célula como ARNm y llevar a cabo directamente la síntesis de proteínas virales. Los "virus +", pueden infectar con el ácido nucleico solo, salvo los retrovirus que siendo +, necesitan una transcriptasa reversa asociada al genoma para poder infectar. Por el contrario, se denomina "virus -" cuando el genoma no puede actuar directamente como mensajeros. El ARN - puede actuar como molde, para la síntesis de ARN +, el ARNm viral.
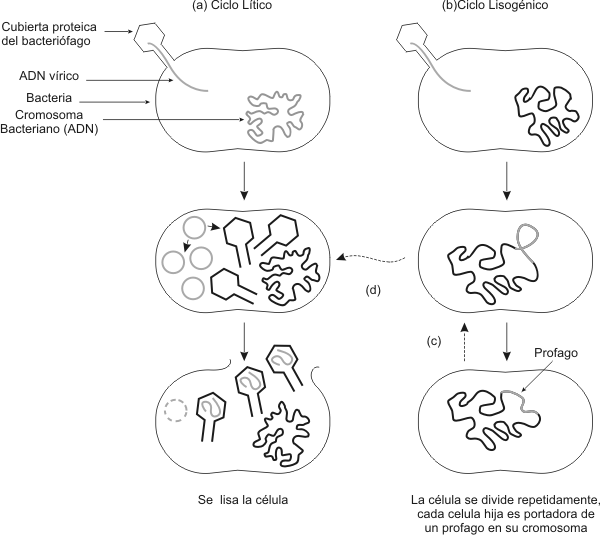
BacteriÓfagos Los Bacteriófagos son virus específicos de las bacterias. Bacteriófagos, significa que se "alimenta" o multiplica a expensas de bacterias. Los Bacteriófagos que infectan células huésped, pueden establecer dos tipos de procesos: 1- Ciclo Lítico: en este tipo de ciclo el virus produce inmediatamente gran cantidad de ácidos nucleicos virales y proteínas de la cápside. Estos se ensamblan produciendo nuevas partículas virales que son liberadas al medio al producirse la lisis celular. 2- Ciclo Lisogénico: en este ciclo la relación entre célula huésped y virus, puede prolongase por periodos variables de tiempo. El virus integra su genoma al cromosoma bacteriano, replicándose conjuntamente el ácido nucleico del parásito y el del huésped. Un virus bacteriano integrado al cromosoma se denomina profago. Por lo tanto el profago se replica junto con el ADN bacteriano. En determinadas circunstancias (por ejemplo ruptura del ADN bacteriano por luz ultravioleta o agentes químicos), el profago se activa, y comienza la producción de ácido nucleico viral y proteínas virales, produciendo luego la lisis celular. Las bacterias que portan profagos se denominan lisogénicas. Los Bacteriófagos que pueden integrarse como profagos y que no lisan inmediatamente a las células se denominan fagos atenuados.
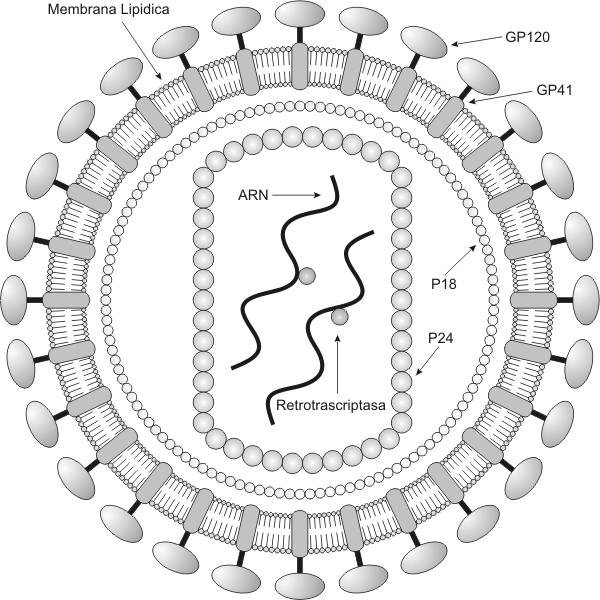
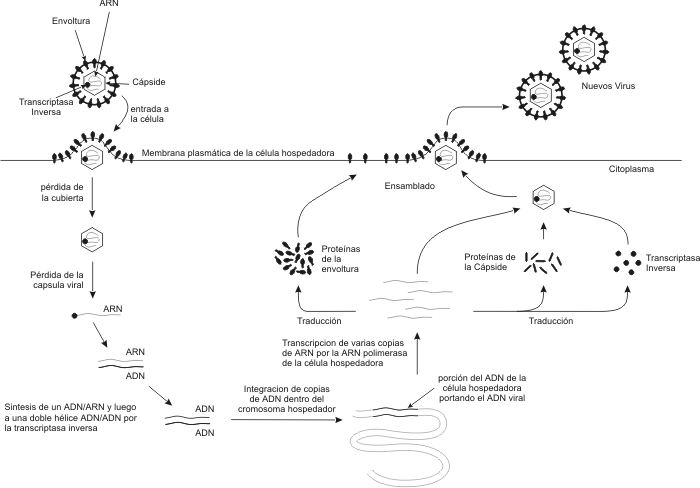
Los virus como vectores Los virus pueden servir como vehículos (vectores), para transferir material genético de una célula a otra. Este fenómeno se denomina transducción. Durante el ciclo lítico, el ADN del huésped queda fragmentado y alguno de esos fragmentos puede ser tomado al azar y quedar dentro de la cápside. De esta forma, cuando el virus infecta una nueva célula, transporta genes de una antigua célula huésped a otra nueva. Virus de la Inmunodeficiencia Humana (HIV) El SIDA (síndrome de inmunodeficiencia humana), es una enfermedad infecciosa crónica producida por el virus HIV. Esta enfermedad esta caracterizada por una deficiencia inmunológica progresiva, con la expresión clínica de infecciones oportunistas y/o tumores. El HIV es un retrovirus. Los retrovirus son virus cuyo material genético es ARN. Una vez que el virus ha penetrado en una célula, una enzima llamada retrotranscriptasa, produce ADN a partir del ARN viral. El ADN recién sintetizado viaja al núcleo y se integra al ADN cromosómico . En este punto la infección se ha hecho permanente , y la forma integrada del virus se denomina provirus. El HIV es un LENTIVIRUS. Por lo tanto su ciclo vital intracelular (ciclo de infección) puede prolongarse por años. Sus genomas son complejos y sus mecanismos regulatorios son solo parcialmente conocidos. Los Lentivirus causan infecciones crónicas. El sistema inmune del hombre reconoce las proteínas del HIV como antígenos y produce anticuerpos contra ellas. Por lo tanto una persona infectada tendrá circulando en sangre anticuerpos contra las proteínas del HIV. En este aspecto se basa el test de diagnóstico más utilizado en la actualidad: el test de ELISA (inmunoensayo ligado a enzima).
El virión del HIV, esta recubierto por una membrana lipídica, por lo tanto se trata de un virus envuelto. De la membrana sobresalen glicoproteínas: la gp41 y la gp120. La membrana compuesta por lípidos y proteínas recubre el núcleo (core) del virión, formado por las proteínas p28 y p24. En el core se encuentran el ARN del virus y la enzima transcriptasa inversa. Ciclo Vital Intracelular de un retrovirus Empieza cuando un virión o partícula vírica , se une a la superficie externa de una célula susceptible. Este primer estadio del ciclo se lo denomina adsorción. Luego el virus fusiona su envoltura lipoproteica con la membrana celular, introduciendo en la célula su nucleocápside junto con el ARN que constituye su dotación genética. En cada partícula viral se encuentran dos cadenas de ARN vírico. A este proceso se lo conoce como penetración. La enzima transcriptasa inversa es una ADN polimerasa que primero produce una copia de ADN simple cadena que a continuación se copia a si misma obteniéndose ADN doble cadena. Por lo tanto este ADN doble cadena se obtuvo a partir de ARN. La síntesis del ADN doble cadena ocurre junto con la degradación del ARN original. El ADN doble cadena (provirus), emigra hacia el núcleo y se integra en el propio ADN celular. La integración de este ADN doble cadena en el cromosoma del huésped es necesaria para la síntesis de nuevas moléculas de ARN, por la ARN polimerasa celular, ya que el virus carece completamente de la maquinaria metabólica necesaria para realizar la transcripción y la síntesis de proteínas necesarias para la cápside y la misma transcriptasa reversa.
Virus de la Hepatitis B (VHB) La hepatitis B en si misma, es un problema sanitario grave y muy extendido . Pero encierra una amenaza peor. El virus que la produce es el carcinógeno humano más importante después del tabaco. Trescientos millones de personas , la mayoría habitantes con escasos recursos asistenciales, están crónicamente infectados con el virus y tienen una probabilidad muy elevada de contraer cáncer de hígado. En el tercer mundo, el virus suele transmitirse de madre a hijo, durante el primer mes de vida, y principalmente durante el nacimiento. Si el pequeño es niña, se convertirá probablemente en portadora crónica y transmitirá el virus de la hepatitis B (VHB) a su descendencia cuando alcance la edad fértil. En cambio en los países desarrollados su incidencia es mayor en adultos, que por su profesión tienen contacto directo con la sangre (cirujanos, enfermeras, dentistas), los receptores de sangre u hemoderivados o personas que reciben tratamientos de diálisis o drogadictos intravenosos. El virus de la hepatitis B (VHB) es mucho más contagioso que el HIV.
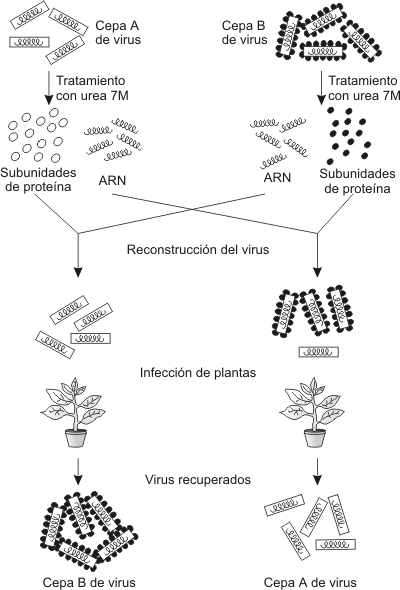
Experimento de Frenkel-Conrat y Singer
Este experimento demostró que el ARN es el material genético del virus del mosaico del tabaco. Los resultados del experimento son extensibles a otros tipos de virus con genoma a ADN. Viroides Los viroides son agentes infecciosos que poseen al igual que los virus un solo tipo de ácido nucleico y son parásitos absolutos, pero y esta es la gran diferencia con los virus, carecen de cápside y envoltura. Por lo tanto los viroides están constituidos solo por una secuencia de nucleótidos, además los viroides carecen de información para la síntesis de proteínas, en cambio los virus siempre poseen dicha información. Priones Las partículas infecciosas llamadas priones, están constituidas únicamente por una proteína de aproximadamente 250 aminoácidos. Es decir carecen completamente de ácidos nucleicos. Es esta la razón por la cual fue resistida durante mucho tiempo, la hipótesis de que las proteínas por si solas podían ser la causa de enfermedades infecciosas. De acuerdo al dogma imperante hasta 1980, las enfermedades transmisibles (infecciosas) necesitaban material genético, para que la infección se asentara en el huésped. Hasta ese momento eran los virus los agentes infecciosos más pequeños conocidos, y todos ellos poseen ADN o ARN como material genético necesario para codificar sus proteínas y dirigir la replicación viral en el huésped. Pero ahora sabemos que las partículas proteínicas infecciosas (priones), pueden ser el sustrato de diversas enfermedades, hereditarias o contagiosas. Este comportamiento dual tanto infeccioso como hereditario era desconocido. Posteriormente se descubrió que los priones se multiplican por una vía increíble y desconocida hasta ese momento: convierten proteínas normales en MOLECULAS INFECCIOSAS, con solo alterar la estructura proteica. Las encefalopatías espongiformes transmisibles (EET), son las enfermedades degenerativas del sistema nervioso central que afectan a animales y seres humanos causadas por los priones. Se denominan espongiformes ya que el cerebro adquiere un aspecto parecido al de una esponja. Las EET que sufren los seres humanos son el Kuru (o muerte de la risa), la enfermedad de Creutzfeldt -Jakob (ECJ), el síndrome de Gerstman-Straussler-Scheinker (GSS) y el insomnio Familiar Fatal (IFF); las EET de animales, incluyen el scrapie (del ingles to scrape, raspar, por la tendencia de los animales infectados a rasparse contra postes , troncos o cercas para combatir la picazón) de ovejas y cabras, la enfermedad de agotamiento crónico de mulas y ciervos en cautiverio y la encefalitis espongiforme bovina (EEB), o enfermedad de la vaca loca. Las EET se caracterizan por su prolongado periodo de incubación (en el hombre puede tener un periodo de incubación de 30 o mas años), generalmente asociadas a declives progresivos de las funciones motoras y cognitivas (enfermedad activa), y por su evolución inevitablemente fatal. Las EET en el ser humano generalmente aparecen en personas de edad avanzada. Aparentemente todas las EET son causadas por el cambio en la estructura de una proteína normalmente presente en las membranas celulares, denominada proteína priónica (PrP). La forma anormal de la PrP se designa PrPsc (scrapie), para diferenciarla de la forma normal llamada PrPc (celular). La secuencia de aminoácidos (estructura primaria) de la PrPc y la PrPsc es idéntica lo que varia es su conformación (plegamiento en el espacio). De acuerdo a esta teoría la proteína alterada (PrPsc), puede unirse a la proteína normal (PrPc) y cambiar su conformación, transformándola a su vez en una proteína alterada. De esta forma se propagaría la enfermedad y se generarían nuevas proteínas infecciosas. De esta forma el pasaje de la forma normal a la patológica es catalizada por el mismo prión (PrPsc), por lo tanto solo hace falta una pequeña cantidad de este para provocar la transformación de toda la proteína normal, ya que se trata de un fenómeno de crecimiento exponencial. Recientemente , se ha aceptado que los priones pueden ser transmitidos, posiblemente por comida, inoculación directa en el cerebro, piel, músculo o estómago. Por esto la epidemia de EEB, ocurrida en Gran Bretaña provoco un enorme interés en todo el mundo. Desafortunadamente han aparecido en ese país una nueva variedad de la ECJ (casos en personas mucho mas jóvenes que lo usual), lo que probaría una relación causal entre la EEB y los casos seres humanos. El gobierno británico tuvo que admitir la posibilidad de que la aparición de estos extraños casos de la ECJ hubieran sido provocados al ingerirse carne vacuna infectada (tejido nervioso). Al principio habíamos hablado de la dualidad de los priones, por un lado partículas infecciosas y por el otro responsables de enfermedades hereditarias. Esto es naturalmente confuso. Por ejemplo, ciertas enfermedades priónicas como la GSS, son hereditarias. Esta enfermedad tiene una herencia autosomal y dominante, lo cual significa que si un padre desarrolla la enfermedad , los hijos tienen un 50 por ciento de probabilidades de desarrollarla. La explicación a estos hechos vino dado por el descubrimiento de mutaciones génicas puntuales en la secuencia de nucleótidos del gen que codifica la PrP . Estos genes mutados provocan cambios en la secuencia de aminoácidos de la proteína PrP. Estos cambios podrían incrementar la probabilidad de la transformación de la proteína PrP mutante de una forma normal a una anormal patógena. Diferentes mutaciones en el gen provocarían diferentes proteínas mutantes, con mayor o menor tendencia a transformarse en la forma aberrante patógena. Esto explica también las distintas enfermedades priónicas hereditarias, la ECJ esporádica , un 15 por ciento de los casos hereditaria y la GSS autosomal dominante.
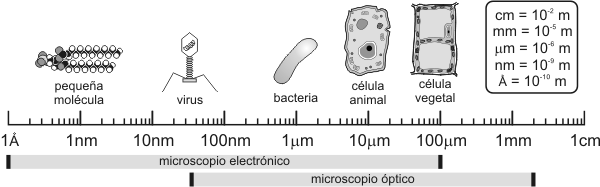
TÉCNICAS DE ESTUDIO DE LA CELULA Una combinación de métodos ayuda al estudio de la célula El desarrollo de la biología celular y molecular se produce en forma paralela a la invención de instrumentos y técnicas biofísicas y bioquímicas que extienden los sentidos a nuevos límites y acrecientan el conocimiento de la estructura y funcionamiento de la célula. El acceso a este tipo de conocimiento resulta dificultoso pues las células son pequeñas y transparentes. El ojo humano sólo puede discriminar dos puntos separados por más de 0,1 mm ó 100 micrómetros (mm). La mayoría de las células son mucho más pequeñas y se necesita del microscopio óptico cuyo límite de resolución es de 0,2 mm. Para estructuras más pequeñas, que midan entre 0,4 y 200 nanómetros (nm), se requiere del microscopio electrónico. Para mayores detalles, consulte la Tabla 1a.2 Como se aprecia, las principales unidades de medida que se emplean corrientemente en microscopia no son las de uso cotidiano. En la siguiente tabla se expresan aquellas unidades y sus equivalencias:
Por lo tanto: 1m=102 cm=102 mm=103 mm=106 nm=109 Por otra parte, la mayoría de los componentes de la célula viva son transparentes debido a su alto contenido de agua. Al disecarla no presenta un significativo contraste. Entonces la tinción selectiva de ciertos componentes celulares resuelve el problema del contraste. Sin embargo la mayoría de estas técnicas conduce a la muerte de la célula y aún más, a cambios químicos y morfológicos que no se encuentran en las células activas y en la matriz extracelular. Cabe destacar que algunos pigmentos del citoplasma y de organoides vegetales pueden absorber determinadas longitudes de onda (l) [1] y no necesitan de previo tratamiento para su observación al microscopio óptico. Para el estudio de la célula viva se emplean técnicas ópticas especiales como, por ejemplo, la microscopia de contraste de fase o la de interferencia.
En el nivel macromolecular, la difracción de rayos X resulta ser la técnica más adecuada para estudiar la configuración molecular de proteínas, de ácidos nucleicos y de algunos entes prebióticos tales como los virus. Los compuestos químicos que constituyen la célula pueden ser detectados, descriptos y localizados dentro y fuera de la célula mediante métodos adaptados de la Química Analítica. Las técnicas aislamiento y separación específica de células, organelas y macromoléculas y el preparado de cultivos celulares para el estudio detallado son herramientas invalorables para el avance de la biología celular y molecular. Los microscopios proveen ventanas para el mundo de la cÉlula
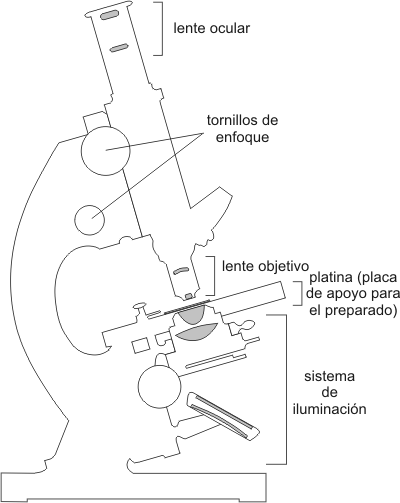
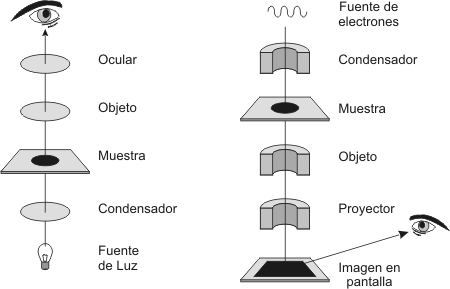
El descubrimiento y estudio temprano progresó con la invención y mejora de los microscopios en el siglo XVII. Los microscopios de varios tipos son aún herramientas indispensables para el estudio de las células. Los microscopios primeramente usados en el Renacimiento tanto como los que son utilizados en los laboratorios hoy, son microscopios ópticos. La luz visible pasa a través de la muestra y de las lentes de vidrio por donde la luz es refractada (“doblada”) de tal manera, que la imagen del espécimen es amplificada cuando se proyecta en el ojo. Dos valores importantes en microscopia son el aumento y el poder de resolución . Entendemos por aumento a las dimensiones aparentes de los objetos comparados con su tamaño real. El poder de resolución es la medida de la nitidez de la imagen; es la capacidad del instrumento para brindar imágenes distintas de dos puntos cercanos. El microscopio óptico nunca puede resolver detalles menores a 0,2mm, la medida de una pequeña bacteria. Esta resolución es limitada por la longitud de onda de la luz visible usada para iluminar la muestra. Los microscopios ópticos o de luz pueden aumentar efectivamente alrededor de 1000 a 1500 veces el tamaño de la muestra real; si se incrementase el aumento la imagen proyectada sería borrosa. La mayoría de las mejoras en microscopia de luz del comienzo de este siglo ha involucrado nuevos métodos para aumentar el contraste. Sin estas técnicas sería imposible para el ojo humano el conocimiento del mundo celular. La mayoría de las organelas son demasiado pequeñas para ser visualizadas por la microscopia de luz. La Biología Celular avanzó rápidamente hace un poco más de cincuenta años con la introducción del microscopio electrónico. En lugar de luz visible, el microscopio electrónico utilizó un haz de electrones [2] a través de la muestra. El poder de resolución está inversamente relacionado con la longitud de onda (l) de la radiación que un microscopio usa y un haz de electrones tiene longitudes de onda mucho más cortas que la luz visible. Para mayores precisiones al respecto, consulte el cuadro 1.4
La mayoría de los microscopios electrónicos modernos pueden lograr una resolución de 0,2 nm, unas 1000 veces mejor que la obtenida con el microscopio óptico. Los biólogos usan el término ultraestructura celular para referirse a la anatomía celular cuando se resuelve por un microscopio electrónico. Hay dos tipos de microscopio electrónico: el microscopio electrónico de transmisión (MET) y el microscopio electrónico de barrido (MEB): · El MET permite develar la ultraestructura de las células y de la matriz extracelular. El haz de electrones es desviado por un campo electromagnético (que actuaría como lente). En este tipo de microscopia se requiere que las muestras tengan un grosor de 100 nm. · El MEB es especialmente útil para estudios de superficie del espécimen. El haz de electrones explora la superficie de la muestra la que usualmente se recubre con una película de oro. El haz excita a los electrones sobre la superficie de la misma muestra y estos electrones secundarios se recolectan y enfocan sobre una pantalla. Esto forma una imagen de la topografía del espécimen. Un importante atributo del MEB es su gran profundidad de campo, la cual resulta en una imagen tridimensional. El ME revela muchas organelas que son imposibles de resolver con MO. Pero el MO ofrece muchas ventajas especialmente para el estudio de la célula viva. Una desventaja de ME es que los métodos químicos y físicos usados para preparar la muestra, no sólo matan a las células sino que puedan introducir artefactos, estructuras peculiares vistas en micrografías que no existe en una célula viva. La preparación de la muestra para el MO y el ME resulta similar si se trabaja con las células muertas. En la tabla 1.9 se comparan los pasos a seguir en ambos tipos de microscopia para la preparación de la muestra:
Las diferencias más notables de los principales componentes del MO y del ME y sus características singulares se especifican en la siguiente tabla.
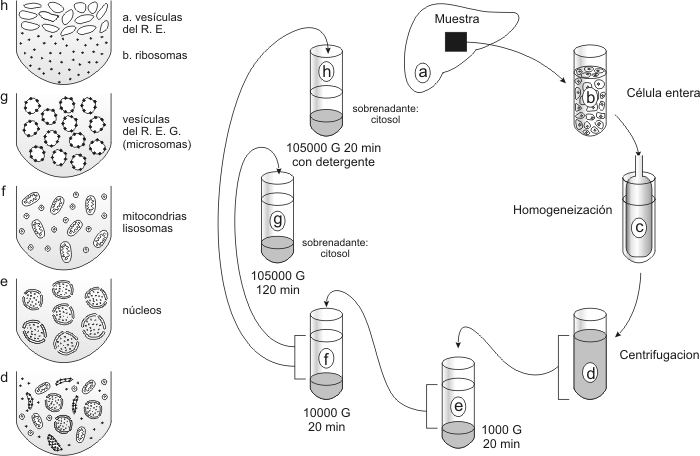
Las cÉlulas vivas pueden ser observadas a travÉs del microscopio Se pueden utilizar ingeniosamente las propiedades ondulatorias de la luz aumentando el contraste de las estructuras transparentes. Así surgieron varias clases de microscopios ópticos convirtiéndose en herramientas destacadas en el estudio de la célula. Más aún, si se considera la posibilidad de que algunos componentes de la célula puedan perderse o distorsionarse durante la preparación de la muestra. En consecuencia, el examen de la célula viva (sin fijación ni congelación) resulta útil. Los microscopio de contraste de fase y de interferencia son sistemas ópticos especiales que logran intensificar la escasa interferencia [3] y proporcionan mayor contraste que el obtenido con un MO común. En el microscopio de campo oscuro, el haz luminoso se proyecta oblicuamente y los sistemas de lentes detectan la luz reflejada por la muestra que se ve como un objeto brillante contra un fondo oscuro, lográndose un gran relieve de rasgos de la célula que son invisibles en el MO. El microscopio de luz polarizada puede proveer datos sobre la estructura a nivel molecular de las células y los tejidos no sólo en las células vivas sino en preparados post-mortem. Ciertos componentes celulares y tisulares se comportan peculiarmente cuando la luz polarizada (luz que gira únicamente en un plano) los atraviesa. Las organelas pueden aislarse para estudiarlas más profundamente La descripción de las diversas organelas dentro de la célula revela poco acerca de su función. La Biología Celular actual desarrolla la integración de la Citología con la Bioquímica, es decir el estudio de la estructura celular junto con el análisis de los procesos químicos de la vida (metabolismo). El fraccionamiento celular ha sido particularmente importante en esta ciencia. El objetivo del fraccionamiento celular es disgregar las células separando las organelas principales de modo que, sus funciones individuales puedan ser estudiadas. El instrumento usado para fraccionar las células es la centrífuga capaz de girar a diversas velocidades. La más poderosa, llamada ultracentrífuga puede dar vueltas tan rápidamente como 80000 revoluciones por minuto (RPM) y aplicar fuerzas sobre las partículas de 500000 veces mayores que la fuerza de la gravedad (500000g).
El fraccionamiento celular comienza con la homogeneización, o sea la ruptura de la célula. El objetivo es romper las células sin dañar severamente sus organelas. El homogenato se centrifuga lográndose la separación de las partes de la célula en dos fracciones: el pellet que consiste en las estructuras más grandes depositadas en el fondo del tubo y el sobrenadante constituido de partes más pequeñas suspendidas en el líquido por encima del pellet. El sobrenadante es transvasado a otro tubo y centrifugado otra vez. El proceso es repetido incrementando la velocidad en cada paso. De esta manera se consigue recolectar componentes más pequeños de los sucesivos pellets. Los detalles de esta marcha se aprecian en la Fig.1.19. El fraccionamiento celular permite preparar los componentes específicos de la célula tanto cualitativa como cuantitativamente. Las poblaciones celulares pueden ser separadas por el citÓmetro de flujo Este instrumento posee un tubo muy delgado por donde se hacen pasar a las células una tras otra. Un determinado tipo celular emitirá fluorescencia, previo tratamiento con fluorocromo (pigmento fluorescente). Esto permite la discriminación y, por lo tanto, la separación de las células así tratadas de acuerdo a la fluorescencia que emitan. Las cÉlulas vivas pueden ser estudiadas mediante el cultivo celular Con el objetivo de preservar toda la información que sea posible sobre todos los tipos celulares, se han desarrollado técnicas de disociación de las células. En los primeros tiempos, la técnica consistía en colocar un explante de un pequeño trozo de tejido en un medio compuesto por suero sanguíneo, extracto de embriones y solución salina. Hoy se conocen los requerimientos nutricios de las células eucariontes. Se las mantienen y hacen crecer en un medio sintético enriquecido de suero. Se distinguen tres tipos de cultivos: · Cultivos Primarios se obtienen de los animales o vegetales. El tejido se separa en condiciones asépticas y se corta en pequeños trozos y se los trata con tripsina (enzima proteolítica) que disocia los agregados celulares y genera una suspensión de células libres que se cultivan en un medio apropiado. · Cultivos Secundarios se obtienen mediante el tratamiento con tripsina de un cultivo primario, seguido de un nuevo cultivo en una caja de Petri. · Cultivos de líneas establecidas cuyo crecimiento se prolonga debido a las condiciones cancerosas de las células. Entre las más usadas se encuentran las células HeLa. A pesar de tales anomalías son muy útiles para el estudio del cáncer.
Las molÉculas orgÁnicas son estudiadas dentro y fuera de la cÉlula A nivel molecular, los organismos están formados por relativamente pocos tipos de compuestos. El agua constituye alrededor del 70% de la mayoría de los seres vivos. De los demás compuestos, los principales son los glúcidos, los lípidos, las proteínas y los nucleótidos, muchos de ellos combinados para formar los ácidos nucleicos, ácido ribonucleicos (ARN) y ácido desoxirribonucleico(ADN). Aunque hay muchos otros compuestos presentes, estas cuatro categorías constituyen la mayoría de las moléculas orgánicas de los seres vivos y son los actores de los procesos metabólicos. Por lo tanto, su localización y función dentro de la célula como un estudio más detallado fuera de ella se hace necesario. Algunos ejemplos de técnicas para el estudio, la ubicación y la cuantificación de moléculas orgánicas se mencionan a continuación: ReconstrucciÓn de imÁgenes a partir de electromicrografÍas Las microfotografías electrónicas poco claras de cristales de moléculas o estructuras supramoleculares se colocan en la trayectoria de un haz de rayos láser para obtener un diagrama de difracción óptica. Este diagrama es procesado por un programa de computación consiguiendo la reconstrucción de las moléculas individuales en las fases y amplitudes correspondientes a un área de la microfotografía. DifracciÓn de rayos X Este método consiste en el bombardeo de la molécula por un delgado haz de rayos X y provoca la dispersión (difracción) de la radiación a través de los electrones de los átomos de la muestra. Este haz disperso debe alcanzar una placa fotográfica. La estructura molecular tridimensional se deduce de las posiciones y las intensidades de las manchas registradas en la placa fotográfica. Los mÉtodos citoquÍmicos Los compuestos químicos se identifican "in situ" por medio de métodos citoquímicos. Este objetivo es cualitativo y cuantitativo y muchas veces persigue el estudio de los cambios dinámicos producidos en el contenido químico celular en las distintas etapas del metabolismo. Para ello, el compuesto químico debe ser: a) inmovilizado en posición original e, b) identificado por un procedimiento específico para el compuesto o para un grupo químico característico que posea. Los métodos físicos y reacciones químicas similares a las implementadas en Química Analítica pero adaptadas a tejidos se usan para llevar a cabo este tipo de identificación. Las diversas molÉculas de la cÉlula pueden localizarse por las tÉcnicas inmunohistoquÍmicas Cuando los vertebrados superiores, los animales e incluso el hombre, son expuestos a la actividad de los microorganismos o a moléculas ajenas a sus ultraestructuras, ya sea naturalmente o por inyección, aquellos sintetizan anticuerpos. Estos son proteínas que pueden fijarse selectivamente a los materiales extraños que desencadenaron su síntesis. Por lo tanto, el organismo produce tantos anticuerpos como partículas extrañas (antígenos) son reconocidas. Así, se puede obtener una gran variedad de anticuerpos. Los anticuerpos son herramientas experimentales muy versátiles para reconocer y manipular células y moléculas. Si un animal de una especie determinada es inyectado con un componente celular o molecular de otra especie, el primero logra fabricar anticuerpos capaces de reconocer y fijar fuertemente aquel componente (el del segundo animal). A estos anticuerpos se acoplan colorantes fluorescentes que son visibles al MO. De modo similar, con el M.E. se pueden detectar moléculas trazadoras acopladas en anticuerpos. De esta manera, los componentes celulares y moleculares quedan “teñidos” en forma diferencial por los anticuerpos que se fijan a ellos. Como todavía no se han presentado los aspectos fundamentales de las moléculas orgánicas, otros métodos fisicoquímicos de más fina determinación, purificación y cuantificación de las biomoléculas se mencionarán en las próximas unidades. Como se verá, dichas técnicas son de uso corriente en el diagnóstico médico rutinario. CUESTIONARIO DE AUTOEVALUCIÓN 1. ¿Cuál de los siguientes organoides no forman parte del Sistema de Endomembranas?: a. envoltura nuclear b. cloroplasto c. aparato de Golgi d. retículo endoplasmático
2. Un determinado veneno destruye el citoesqueleto. ¿Cuál de las siguientes funciones sería afectada directamente?: a. la división celular b. la respiración celular c. la fotosíntesis d. la síntesis de proteínas
3. ¿Cuál de las siguientes organelas está presente en la célula animal y vegetal?: a. cloroplasto b. pared celular c. mitocondria d. centríolos.
4. ¿Qué organela está presente en una célula procarionte?: a. mitocondria b. ribosoma c. cloroplasto d. retículo endoplasmático
5. ¿En qué parte de la mitocondria aparecen las crestas? 6. ¿Qué organela carece de membrana? 7. ¿En qué parte de la bacteria se realiza la respiración? 8. Si Ud. tuviera que argumentar acerca de que los virus son organismos vivos, ¿qué características estructurales y funcionales del virus podría utilizar para apoyar su argumentación? 9. ¿Qué tipo de células pensaría Ud. que alcanzarían el mayor tamaño: una célula muy aplanada o una esférica? ¿Por qué? 10. ¿Por qué casi todas las células casi siempre son microscópicas? 11. ¿Cuáles son las propiedades fundamentales que comparten todas las células? Describa la importancia de cada una de ellas. 12. Compare en un cuadro las diferencias estructurales, funcionales y metabólicas entre las células procariontes y eucariontes. 13. ¿Qué propiedades distinguen a un virus de una bacteria? 14. Enumere los pasos de la infección viral, y describa brevemente cada paso. 15. Utilizando el Sistema Métrico Decimal resuelva las equivalencias en metros de: a) 1 nm; b) 1 mm. 16. Indique el poder de resolución del: a) M.O.; b)M.E.T.; c)M.E.B. 17. ¿Cuántas veces son mayores los aumentos del M.E. en relación a los del M.O.? 18. ¿A qué es igual el poder de resolución?: a. AN / 0.61 x l b. 0.61 x l / AN c. 0.61 / AN x l d. 0.61 x AN /l.
19. ¿Qué compuesto se usa como fijador en microscopia electrónica? ¿Y en microscopia óptica? 20. Explique qué técnicas utilizaría para crear suficiente contraste cuando se usa el M.O. y el M.E. 21. ¿Qué tipo de microscopios se usan para la observación de la células vivas? 22. ¿Cuáles son las ventajas de estudiar las células con el M.E.T. y el M.E.B.? 23. ¿Cuándo utilizaría la microscopia de contraste de fase? ¿Por qué? 24. ¿Para qué se utiliza la técnica de fraccionamiento celular? ¿Qué se obtiene en cada paso? 25. ¿Qué es un anticuerpo? ¿Por qué es una herramienta útil en la investigación biológica? 26. ¿Para qué se utiliza la difracción de rayos X? 27. Cuando una célula de forma esférica crece, los mm2 de superficie de la membrana plasmática por mm3 del volumen celular: a. decrece b. aumenta c. se mantiene constante d. se vuelve más delgada
28. El microscopio de interferencia es: a. una variante del M.O. b. una variante del M.E. c. útil cuando se colorea la muestra d. utilizado para visualizar átomos
29. La ultracentrífuga se emplea en: a. citometría de flujo b. microscopia electrónica c. fraccionamiento celular d. difracción de rayos
30. Las muestras de células vivas o preparados fijados se pueden estudiar indistintamente con: a. microscopia electrónica b. microscopia de interferencia c. microscopia de luz polarizada d. difracción de rayos X.
BIBLIOGRAFÍA l Alberts, B et al; (1996) Biología Molecular de la Célula. 3ra Edición. Ediciones Omega S.A. Barcelona. l Brock, T; (1997) Biology the Microorganisms. 8th Edition. Prentice Hall. NY l Campbell, N; (1997) Biology. 4th Edition. the Benjamin Cummings Publishing Company. Inc. California l Castiñeira de Dios L. y col. (1999). Cuadernillos de Biología e Introducción a la Biología Celular Nº 2. Bs.As. Ediciones CCC-Educando. l Curtis y Barnes (1992). Biología. 5ª Ed. Bs.As. Editorial Médica Panamericana. l De Robertis(h); Hib; Ponzio. (1996).Biología Celular y Molecular de De Robertis. 12º Edición. El Ateneo. Bs.As. l De Robertis, E.; Hib, J.; (1998) .Fundamentos de Biología Celular y Molecular. El Ateneo. Bs.As. l Freidfelder, D. (1982) Physical Biochemestry. 2ª Ed. W.H. Freeman and Co. l Holtzman, E. y Novikoff, A.B.(1986). Estructura y Dinámica Celular. 3ª Ed. México. Interamericana. l Karp, G.; (1998) Biología Celular y Molecular. Ed. Mc Graw Hill Interamericana. México. l Moreno Azorero, R. y Schwartzman, G.; (1986). Principios de Biología Celular. El Ateneo. Bs.As. l Smith and Wood; (1997) . Moléculas Biológicas. Ed.Addison-Wesley, Iberoamericana S.A. l
Solomon
y col. (1998) . Biología de Villee. 4ª. Ed. Mex. McGraw-Hill. Interamericana [1] La longitud de onda (l) es la distancia entre dos puntos consecutivos que vibran en fase. Siempre que dos puntos estén separados por una distancia igual a la que recorre la onda en un período, o a un múltiplo de ella, los dos puntos vibrarán en fase. [2] Los electrones pueden considerarse como una variedad de radiación de pequeña longitud de onda. [3] Interferencia: Es un fenómeno físico producido entre ondas cuyos "picos" y "valles" no coinciden, es decir no están en fase. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
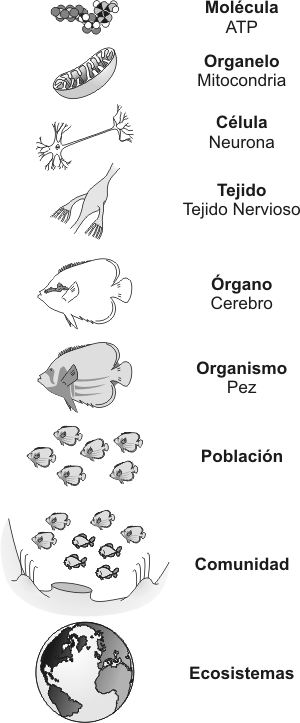
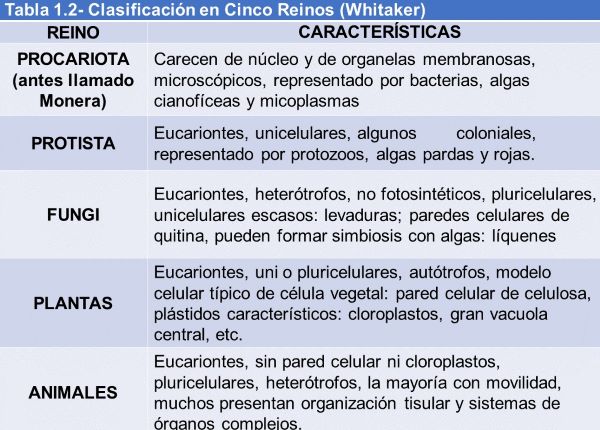
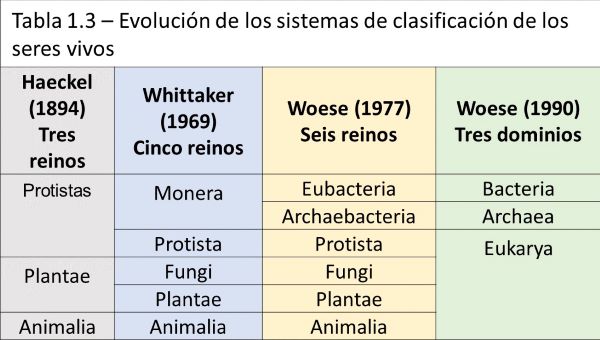
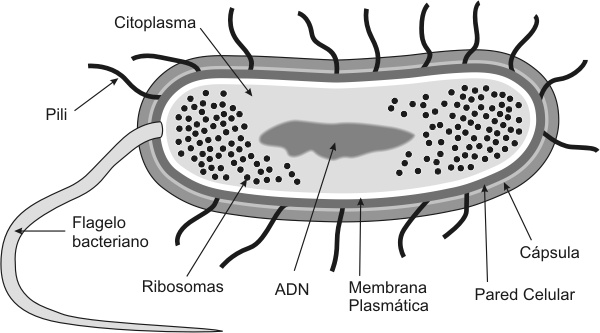
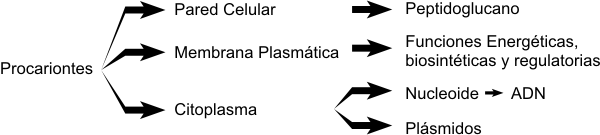
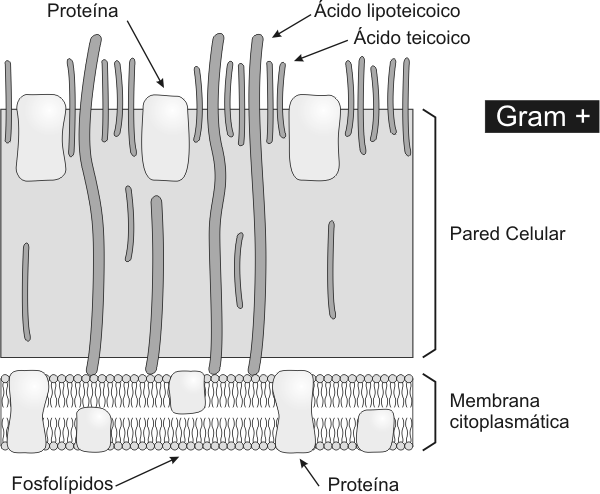
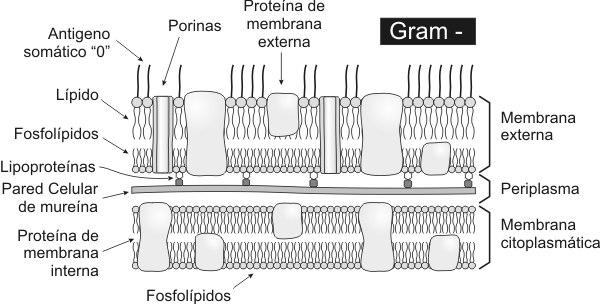
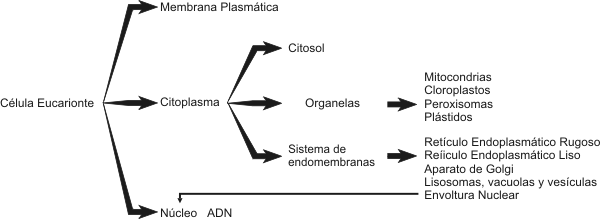
 Fig.1.5- Esquema de la ultraestructura tridimencional de una célula animal y sus principales componenetes
Fig.1.5- Esquema de la ultraestructura tridimencional de una célula animal y sus principales componenetes