MEMBRANA PLASMÁTICA
|
ISBN
Nº 987-4-4580-2
|
Introducción
Las células están separadas del medio que las rodea por una delgada lámina denominada membrana plasmática, que define los límites de las mismas.
Hace 3700 millones de años, la formación espontánea de una estructura similar a la membrana plasmática de las células actuales permitió aparición de los primeros seres vivos. Sin esta barrera protectora, las células estarían expuestas a los rigores del mundo externo, no podrían regular su medio interno y, en consecuencia, no serian viables. La membrana plasmática no aísla a la célula completamente sino que constituye una barrera altamente selectiva, que tiene la propiedad de regular el intercambio de materiales entre la célula y el medio que la rodea.
La membrana es una estructura muy delgada: sólo tiene un espesor de 6 a 10 nm (1nm=10-9m). Por lo tanto, se necesitarían mil membranas plasmáticas apiladas, una sobre otra, para igualar el espesor de esta hoja de papel. Precisamente debido a su delgadez, cuando se examina una célula al microscopio óptico convencional, puede observarse sin dificultad el interior de la misma; en el mejor de los casos podrá apreciarse el contorno de la membrana, pero nunca podrá distinguirse su ultraestructura. Recién las primeras microfotografías al microscopio electrónico demostraron que la ultraestructura las membranas era siempre la misma. Esta estructura se denominó unidad de membrana y la misma no sólo es válida para la membrana plasmática, sino para casi todas las membranas celulares.
|
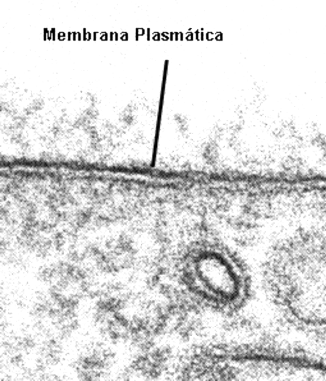
Fig. 4.1 - Microfotografía electrónica de transmisión de una membrana plasmática. Se pueden ver tres líneas paralelas, dos líneas densas alos electrones (2,5 - 3 nm) separada por una capa intermedia clara (3,5-4 nn). Este aspecto conocido como unidad de membrana no es reflejo de una estructura trilaminar a nivel molecular, sino es la expresión de como el osmio, usado como "colorante" se une a la membrana. |
Funciones de la membrana plasmática
Como ya se mencionó, las membranas no son simples barreras sino que:
· Definen la extensión de la célula y establecen sus límites.
· Constituyen barreras selectivamente permeables, dado que impiden el intercambio indiscriminado de sustancias entre el citoplasma y el medio extracelular. La membrana plasmática, gracias a sus propiedades fisicoquímicas, está capacitada para transportar de un lado a otro de la misma determinados solutos, macromoléculas y complejos macromoleculares. Sin embargo, hay moléculas, que a pesar de ser toxicas para la célula, pueden ingresar sin dificultad a la misma a través de la membrana. Un ejemplo seria el CO (monóxido de carbono).
· Controlan las interacciones de la célula con el medio extracelular (tanto con la matriz extracelular como con otras células vecinas). Permite a las células reconocerse, adherirse entre sí cuando sea necesario e intercambiar materiales e información.
· Intervienen en las respuestas a señales externas a la célula. La membrana posee receptores, que son moléculas o conjuntos de moléculas, capaces de reconocer y responder a señales provenientes del medio extracelular portando información especifica. Cuando dichas señales llegan hasta la membrana plasmática, se desencadenan señales internas en la célula, tanto activadoras como inhibitorias de distintos procesos celulares. Como ejemplos de estas señales externas podemos citar a los factores de crecimiento que favorecen la división celular o diversas hormonas como por ejemplo la insulina, que aumenta la síntesis de glucógeno.
Singer y Nicholson propusieron en 1972 un modelo estructural para las membranas al cual denominaron modelo del mosaico fluido. De acuerdo al mismo las membranas son disoluciones bidimensionales de lípidos y proteínas. Según este modelo, la estructura de la membrana sería una delgada lamina formada por dos capas superpuestas de lípidos (también llamadas hemimembranas), con la fluidez propia de los aceites, en la cual se encuentran insertadas proteínas. Esto le confiere el aspecto de un mosaico.
|
Fig. 4.2 -Esquema del "Modelo del mosaico fluido" de las membranas |
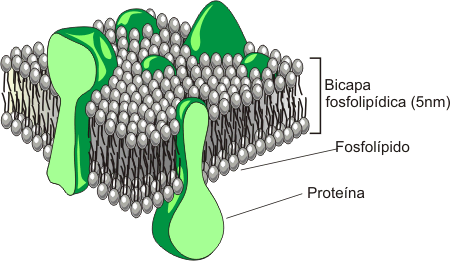
Las membranas no son estructuras estáticas ni rígidas. Están formadas por un conjunto de moléculas hidrofóbicas e hidrofílicas que se mantienen unidas por enlaces, en general, no covalentes. Una de las principales características de las membranas biológicas es su alto grado de fluidez. Esto implica que sus lípidos y proteínas pueden desplazarse libremente en todas las direcciones, pero siempre sobre el plano de la membrana. De allí entonces la denominación de mosaico fluido; a esta propiedad también se la conoce como difusión lateral.
Como puede observarse en el esquema, las membranas también presentan glúcidos unidos por enlaces covalentes a lípidos y proteínas. Esto da lugar a los llamados glucolípidos y glucoproteínas, respectivamente.
Estas membranas carecen de resistencia mecánica y en muchas células, como en el caso de hongos, bacterias y plantas están reforzadas por paredes celulares.
1. COMPOSICIÓN DE LAS MEMBRANAS BIOLÓGICAS
Todas las membranas biológicas de los seres vivos, tanto la membrana plasmática, como las de las organelas, están formadas por:
A. Lípidos
B. Proteínas
C. Glúcidos
La proporción de cada uno de estos componentes varía de acuerdo a la función que realiza cada tipo de membrana. Por ejemplo, las membranas mitocondriales tienen una proporción muy elevada de proteínas (ver Tabla 1).
|
Tabla 1 Composición de las membranas de diferentes células. (Los valores representados como % peso seco de la membrana) |
|||
|
Glóbulos Rojos Humanos |
Staphiloccoccus aureus |
Mielina |
|
|
Lípidos |
30 a 40 |
20 |
60 a 70 |
|
Fosfolípidos |
20 a 25 |
20 |
25 a 30 |
|
Àcido fosfatídico |
< 1 |
- |
0 |
|
Fosfatidiletanolamina |
5 |
- |
5 |
|
Fosfatidilcolina |
7 |
- |
10 |
|
Fosfatidilserina |
5 |
- |
5 |
|
Esfingomielina |
6 |
- |
5 |
|
Cerebrósidos |
<1 |
- |
10 |
|
Colesterol |
12 |
<1 |
15 |
|
Otros Lípidos |
3 |
- |
15 |
|
Proteínas |
60 a 70 |
40 |
20 a 30 |
|
Glúcidos (o restos de glúcidos en glicoproteínas) |
7 |
40 |
Observada en cortes histológicos |
A. Lípidos
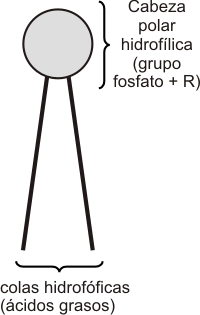
La variedad de lípidos presentes en las membranas es muy amplia; sin embargo, todos poseen una característica en común: son moléculas anfipáticas. Esto significa que sus moléculas contienen una zona hidrofílica o polar y una hidrofóbica o no polar.
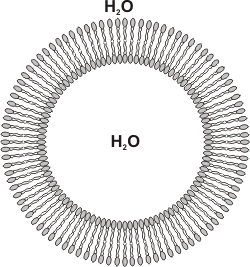
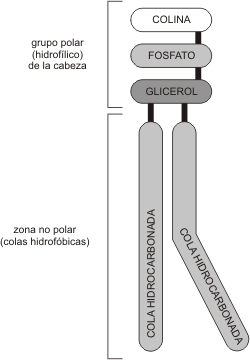
Los fosfolípidos son los lípidos más abundantes en las membranas. Debido a su carácter anfipático, los fosfolípidos, en un medio acuoso se organizan espontáneamente conformando la denominada bicapa lipídica. Las cabezas polares están orientadas hacia el medio acuoso (intra y extracelular) y las colas hidrofóbicas hacia el medio lipídico, es decir, al interior de la bicapa, constituyendo la matriz de la membrana. A su vez, estas bicapas tienden a cerrarse espontáneamente sobre sí mismas formando vesículas, es decir, compartimientos cerrados en toda su extensión tridimensional, similares a una esfera.
|
Fig. 4.3 - Esquema de un fosfolípido. |
Fig. 4.4 - Corte esquemático de una vesícula de fosfolípidos. |
La bicapa de fosfolípidos funciona principalmente como armazón estructural de la membrana y como barrera que impide el pasaje de sustancias hidrosolubles a través de la misma; esto último es debido al carácter fuertemente hidrofóbico de la matriz de la membrana.
Los fosfolípidos más frecuentes de las membranas son la fosfatidiletanolamina, la fosfatidilcolina, la fosfatidilserina y la esfingomielina. Los fosfolípidos de las membranas son DIACILGLICERIDOS.
La estabilidad de las bicapas lipídicas esta dada por:
-
interacciones hidrofóbicas entre las colas hidrocarbonadas.
-
fuerzas de van der Waals entre las colas hidrofóbicas.
-
fuerzas electrostáticas y puentes hidrogeno entre las cabezas polares de los lípidos, ya sea entre ellos mismos y con las moléculas de agua de los medios extra e intracelular.
Como se notará todas estas son uniones débiles (no covalentes) y le confieren simultáneamente estabilidad y fluidez a la membrana.
Las cadenas hidrocarbonadas de los ácidos grasos que forman parte los fosfolípidos (también denominadas colas o grupos acilo), pueden presentarse:
-
saturados (sin dobles enlaces)
-
monoinsaturados (con un único doble enlace)
-
poliinsaturados (más de un doble enlace)
En general, los lípidos de membrana contienen un grupo acilo insaturado y otro saturado en su estructura.
|
Fig. 4.5 -Fosfatidiletanolamina-(fosfolípido de membrana) |
Fig. 4.6 - Esquema de un fosfolípido con una cola saturada y una no saturada. |
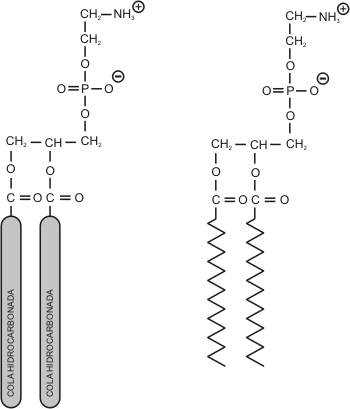
La presencia de ácidos grasos insaturados aumenta la fluidez de la membrana, debido al quiebre de las colas a la altura de los dobles enlaces. Esto impide, o al menos dificulta, que las colas hidrocarbonadas se compacten, restringiendo así las interacciones entre ellas. El hecho de que uno de los grupos acilo de los fosfolípidos esté saturado y el otro no, garantiza una buena fluidez dentro del rango de temperaturas fisiológicas. Por otro lado, cuando las cadenas hidrocarbonadas son cortas, tienen menor superficie para interactuar entre sí; esto último también favorece la fluidez de las membranas.
El colesterol es un esteroide que se encuentra en un alto porcentaje en la membrana plasmática de las células animales. Su concentración varía mucho de un tipo de membrana a otro; en animales hay membranas donde el colesterol constituye hasta el 50% del total de los lípidos. Contrariamente, la mayoría de las células vegetales y bacterianas carecen de colesterol.
El colesterol, al ser también una molécula anfipática, presenta una orientación similar a la de los fosfolípidos: el grupo hidroxilo (polar) se orienta hacia el exterior de la bicapa y el sector hidrofóbico hacia el interior de la misma.
|
Fig. 4.7 - Esquema de la ubicación del colesterol en la membrana plasmática |
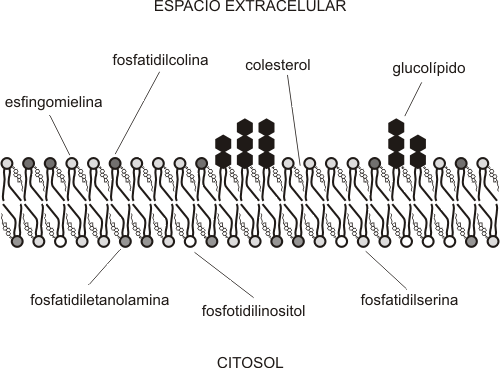
Las funciones del colesterol se pueden resumir de la siguiente mane
- Inmoviliza los primeros carbonos de las cadenas hidrocarbonadas. Esto hace a la membrana menos deformable y menos fluida, es decir, la estabiliza. Sin colesterol, la membrana necesitaría de una pared celular que le otorgue contención mecánica.
- Previene el compactamiento de las cadenas hidrocarbonadas a bajas temperaturas, ya que evita que las colas se junten, aumenten las interacciones débiles entre las mismas y se cristalicen (adopten una estructura muy compacta).
La cardiolipina es un derivado de los fosfolípidos que se encuentra en la membrana interna de la mitocondria. El dolicol es un lípido que se halla en el REG e interviene en la glicosilación de las proteínas.
B. Proteínas (Fig.4.8)
Mientras que los lípidos ejercen principalmente una función estructural, las proteínas no sólo desempeñan un rol estructural sino que además son las responsables de las funciones específicas de las membranas biológicas. Estas según su función pueden agruparse en: enzimáticas, de transporte, receptoras y de reconocimiento. Diferentes membranas tienen distinta proporción y composición de proteínas, de acuerdo a sus funciones. En otras palabras, son justamente las proteínas las que le otorgan distintas funciones a las membranas. Estas en su mayoría son proteínas globulares (estructura terciaria o cuaternaria).
Según su ubicación en la membrana se clasifican en:
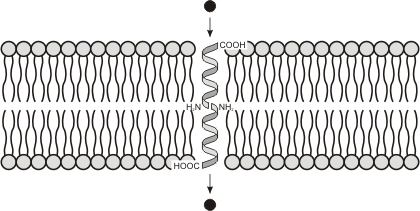
-Proteínas intrínsecas, integrales o transmembrana: Pueden atravesar total o parcialmente la bicapa, asomando a una o ambas superficies de la misma. Únicamente pueden ser extraídas de la membrana por medio de detergentes que rompen la bicapa. Tienen un sector hidrofóbico, que es el que esta insertado en la membrana y una o dos regiones hidrofílicas, expuestas a los medios intra y extracelulares (ambos acuosos). De lo anterior se deduce que estas proteínas son moléculas anfipáticas. La porción que atraviesa la membrana suele presentar una estructura de alfa hélice con una elevada proporción de aminoácidos hidrofóbicos que interaccionan con las colas hidrocarbonadas de la matriz de la membrana. El sector proteico (también llamado dominio) expuesto a los medios acuosos suele tener estructura globular e interacciona con las cabezas polares de los fosfolípidos y con otras moléculas a través de uniones iónicas y puente de hidrógeno.
Dentro de las proteínas integrales encontramos:
-
Proteínas monopaso: La proteína atraviesa una sola vez la membrana.
-
Proteínas multipaso: La cadena polipeptídica atraviesa dos o más veces la bicapa lipídica. Por lo tanto, esta posee varias regiones hidrofóbicas insertadas en la matriz de la membrana alternadas con sectores hidrofílicos que se exponen hacia los medios acuosos.
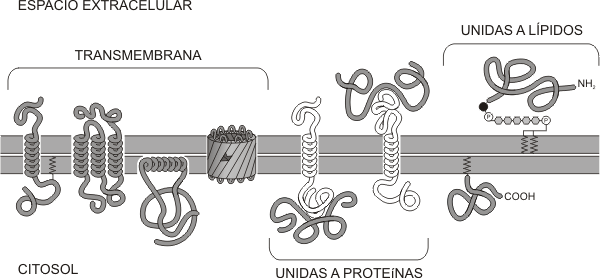
Fig. 4.8 -Asociación de proteínas de membrana con la bicapa lipídica: Transmembrana, atraviesan la membrana como -helice o como láminas plegadas cerradas. Periféricas unidas a proteínas transmembrana por interacciones no covalentes débiles y Periféricas unidas a lípidos mediante uniones covalentes.
Algunas proteínas multipaso atraviesan muchas veces la membrana y forman un cilindro hueco con un interior hidrofílico por el que pueden pasar moléculas pequeñas solubles en agua. Este es el principio de las proteínas canal que se analizaran mas adelante.
Las proteínas integrales pueden difundir lateralmente y rotar sobre su propio eje, pero no pueden realizar movimientos a través del plano de la membrana, o más sencillamente movimiento flip-flop (ver más adelante). Las proteínas integrales suelen desplazarse acompañadas de los lípidos que las rodean ya que estos le ayudan a mantener su conformación.
Sin embargo, algunas proteínas integrales están ancladas a componentes del citoesqueleto y no pueden trasladarse. De esta manera intervienen en la morfología de la célula, por ejemplo alargada (o ahusada), cúbica, cilíndrica, etc.
-Proteínas extrínsecas o periféricas: Se encuentran sobre la cara externa o también interna de la membrana y pueden estar ligadas tanto a las proteínas integrales como a los fosfolípidos por uniones débiles. Se pueden extraer fácilmente con tratamientos no drásticos. Cuando estas se ubican del lado citoplasmático de la membrana suelen interactuar con el citoesqueleto.
C. Hidratos de carbono
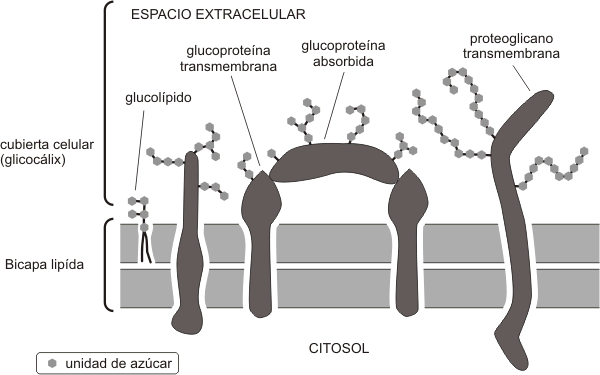
Las membranas celulares contienen entre un 2-10% de glúcidos. Estos se asocian covalentemente a los lípidos (glicolípidos) y a las proteínas (glicoproteínas).
Los glicolípidos (o glucolipidos) presentes en las membranas son los gangliósidos y cerebrósidos. Los gangliósidos se forman por la unión de un oligosacárido con la ceramida. La estructura de los cerebrósidos es similar, sólo que el hidrato de carbono no es un oligosacárido sino una galactosa o una glucosa. (ver capítulo de lípidos)
Los hidratos de carbono de los glucolípidos y las glucoproteínas, en su mayoría oligosacáridos, suelen ubicarse en la cara no citosólica de la membrana plasmática formando una estructura llamada glicocálix (Fig. 4.9 y 4.10), cuyas funciones se pueden resumir de la siguiente manera:
· Proteger a la superficie de la célula de agresiones mecánicas o físicas. Como ejemplo podemos citar a las células situadas en la luz del intestino delgado que presentan un glicocálix muy pronunciado.
· Poseer muchas cargas negativas, que atraen cationes y agua del medido extracelular.
· Intervenir en el reconocimiento y adhesión celular. Actúan como una huella dactilar característica de cada célula, que permite distinguir lo propio de lo ajeno.
· Actuar como receptores de moléculas que provienen del medio extracelular y que traen determinada información para la célula, por ejemplo, receptores de hormonas y neurotransmisores.
|
Fig. 4.9 - Microfotografía electrónica de un glicocalix de epitelio intestinal (izquierda). |
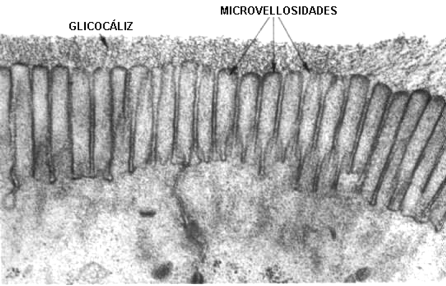
|
Fig.4.10 Esquema del glicocalix de una célula eucariota |
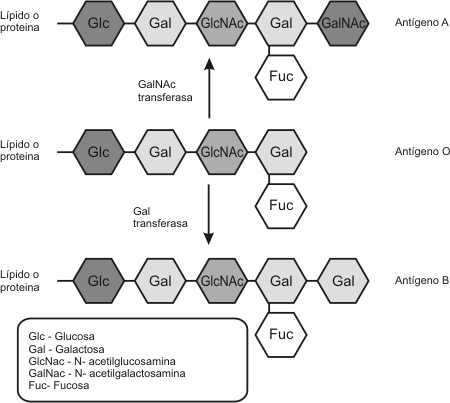
Las diferencias entre los grupos sanguíneos se hallan determinadas por ciertos oligosacáridos muy cortos, presentes en las membranas plasmáticas de los glóbulos rojos o eritrocitos. Estos oligosacáridos sólo difieren en sus monómeros terminales y están ligados a una proteína transmembranosa o a una ceramida de la membrana plasmática. Por ejemplo, los eritrocitos pertenecientes al grupo sanguíneo A, presentan como monosacárido terminal una N-acetilgalactosamina y los del grupo B una galactosa. Cuando ambos monosacáridos terminales están ausentes estamos en presencia del grupo 0 (Fig.4.11).
|
Fig. 4.11 - Grupos sanguíneos |
2. FLUIDEZ DE LA MEMBRANA
Como ya se mencionó, las membranas son estructuras dinámicas donde los componentes pueden desplazarse en todas las direcciones sobre el plano de la bicapa. De ahí que el modelo reciba el nombre de mosaico fluido.
|
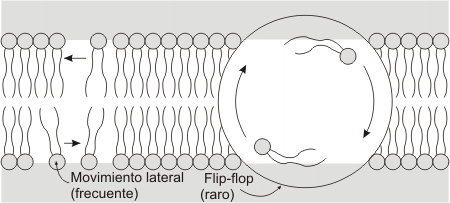
Fig. 4.12 - Movimientos de los fosfolípidos en una bicapa liplídica |
2.1. MOVILIDAD DE LOS COMPONENTES DE LAS MEMBRANAS
Existen tres tipos de movimientos posibles en las membranas:
-
rotación (sobre su propio eje)
-
traslación (o difusión lateral) sobre el plano de la membrana.
-
flip-flop
El movimiento de flip-flop es el intercambio de fosfolípidos de una monocapa (o hemimembrana) a la otra; esta sumamente restringido, debido a la dificultad que posee la cabeza polar para atravesar el medio hidrofóbico de la matriz de la membrana. De allí que no sea un movimiento que ocurra de manera espontánea sino que está mediado por enzimas denominadas flipasas.
Tanto los movimientos de difusión lateral como el de rotación se llevan a cabo sobre la misma hemimembrana de la bicapa lipídica.
2.2. FACTORES QUE AUMENTAN LA FLUIDEZ DE LAS MEMBRANAS
-Ácidos grasos insaturados
-Baja concentración de colesterol
-Altas temperaturas
-Colas hidrocarbonadas cortas (dificultan el empaquetamiento)
Factores que favorecen
la viscosidad
|
Factores que favorecen la fluidez |
|
|
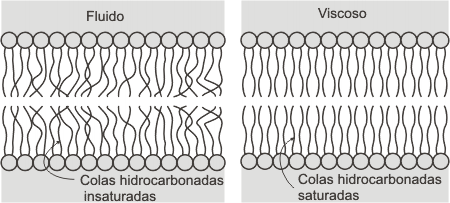 Fig.
4.13 - Esquema de los fosfolípidos de membrana en estado
viscoso y fluído. Fig.
4.13 - Esquema de los fosfolípidos de membrana en estado
viscoso y fluído. |
2.3. EFECTO DE LA TEMPERATURA SOBRE LA FLUIDEZ
El ascenso de la temperatura aumenta la energía cinética entre las moléculas y, por lo tanto, el movimiento de las colas hidrocarbonadas. Esto lleva a una disminución de las interacciones atractivas entre las mismos y a un aumento de los movimientos de rotación y de difusión lateral. Por el contrario, una disminución de la temperatura vuelve más rígida a la membrana ya empaqueta las colas hidrofóbicas de los fosfolípidos e impide sus movimientos. Si la temperatura desciende significativamente, la membrana puede llegar a cristalizarse, con la pérdida consiguiente de muchas funciones vitales de la membrana.
Los organismos que habitan regiones donde hay grandes amplitudes térmicas estacionales varían la composición de los fosfolípidos de sus membranas en forma periódica, asegurando así una fluidez más o menos constante durante todo el año. Por otra parte, organismos que habitan ambientes extremos poseen composiciones fosfolipídicas muy particulares en sus membranas, por ejemplo, los que viven a temperaturas inferiores a los 0ºC tienen membranas muy ricas en lípidos poliinsaturados.
2.4.. DETERMINACIÓN DE LA FLUIDEZ DE LA BICAPA LIPÍDICA
La fluidez de la membrana se pudo determinar experimentalmente tratando células con anticuerpos fluorescentes que eran reconocidos y se unían a las proteínas (receptores) presentes en la membrana plasmática. Gracias a esta técnica, se pudo observar, a través del microscopio, el desplazamiento de los receptores sobre la superficie de la membrana y su agrupamiento en un polo de la célula, donde posteriormente ingresaban por endocitosis (internalización a la célula, ver más adelante).
3. ASIMETRIA DE MEMBRANA
En ambas caras de la bicapa (también denominadas hemimembranas o monocapas) no se encuentran los mismos tipos de fosfolípidos. Si bien estos en su mayoría se sintetizan en la cara citosolica del retículo endoplasmático liso, luego, por medio de movimientos del tipo flip-flop (únicamente permitidos en el REL, gracias a la presencia de flipasas), se van ubicando del lado de la bicapa que les corresponda. Por ej., la fosfatidilcolina y la esfingomielina predominan en la cara no citosolica de la membrana (Fig. 4.7).
La asimetría estructural de las membranas suele manifestarse a través de una asimetría funcional. Esto significa que las funciones presentes en la cara citosolica no son las mismas que aparecen en la cara no citosólica. Por ejemplo, en el caso de la membrana plasmática, las moléculas que intervienen en el reconocimiento celular se ubican casi exclusivamente en la cara expuesta hacia el medio extracelular, pues no tendría mucho sentido que dichas moléculas estuviesen expuestas hacia el citoplasma.
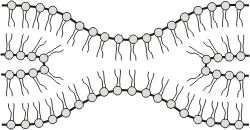
4. FUSIÓN DE MEMBRANAS
Las membranas tienen una elevada capacidad para fusionarse entre sí. Por ejemplo, cuando una vesícula se aproxima a la membrana plasmática, a una cisterna o, inclusive, a otra vesícula, al entrar en contacto ambas superficies, las dos membranas se fusionan, constituyendo a partir de ese momento una sola membrana. Este fenómeno explica el transito de sustancias desde un compartimiento celular a otro, y desde las endomembranas a la membrana plasmática. (ver más adelante endo y exocitosis). Este es el principio en el que se basa la administración de fármacos vehiculizadas dentro de liposomas, que son vesículas fosfolipídicas artificiales que contienen alguna droga de interés terapéutico. Cuando el liposoma se aproxima a la célula blanco (o target), la membrana del liposoma se fusiona con la membrana plasmática liberando su contenido directamente en el citoplasma de la célula. Este fenómeno permite que el contenido del liposoma sólo sea captado por ciertos tipos celulares y no por otros. Técnicas basadas en esta propiedad de las membranas se utilizan, por ejemplo, para combatir células tumorales.
|
Fig. 4.14 -Fusión de dos membranas |
5. PERMEABILIDAD DE LAS MEMBRANAS CELULARES
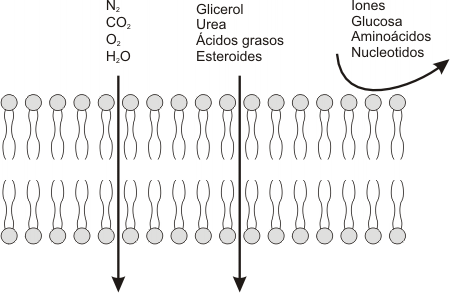
Como ya se ha mencionado la membrana plasmática es una barrera con permeabilidad selectiva que regula el intercambio de sustancias entre el citoplasma y el medio extracelular. Sus propiedades aseguran que las sustancias esenciales, como la glucosa, los aminoácidos y los lípidos entren a la célula fácilmente, que los intermediarios metabólicos permanezcan en la célula y que los productos de desecho, como la urea, abandonen la misma. Todo esto permite a la célula mantener el medio interno relativamente constante. La membrana, debido a sus características hidrofóbicas, es impermeable a la mayor parte de las moléculas hidrosolubles, como la glucosa, los aminoácidos y los iones en general. En cambio, las moléculas hidrofóbicas, siempre y cuando su tamaño no sea demasiado grande, pueden atravesarla fácilmente.
Podemos observar en la figura 4.15, que únicamente atravesarán la membrana las moléculas no polares y pequeñas como el O2, CO2, N2 e incluso el CO (tóxico), compuestos liposolubles como los ácidos grasos y esteroides y, además, a pesar de ser moléculas polares, el glicerol, la urea y el agua. El resto de las moléculas se transfiere de un lado a otro de la membrana gracias a proteínas integrales que actúan como transportadores; sin estos transportadores dichas moléculas no pueden difundir a través de las membranas.
|
Fig. 4.15 - Permeabilidad de la membrana a los diferentes solutos |
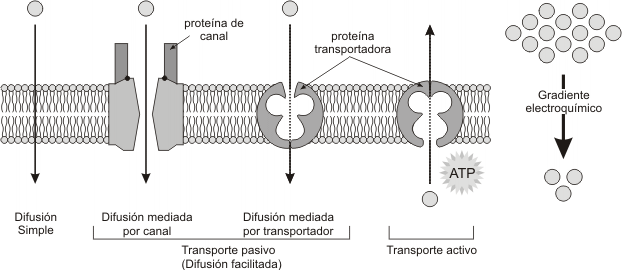
6. MECANISMOS DE TRANSPORTE TRANSMEMBRANA
|
Fig. 4.16 - Distintos mecanismos y estructuras utilizados por los solutos para atravesar las mem-branas de una célula. |
Antes de continuar con los mecanismos de transporte es preciso hacer una breve aclaración acerca del fenómeno de difusión. Si colocamos un soluto en un solvente, las moléculas de soluto, debido a la energía cinética de las moléculas presentes en la solución, difundirán desde la zona donde se encuentran en mayor concentración hacia la zona donde se hallan en menor concentración. Al cabo de un tiempo toda la solución presentará la misma concentración de soluto. Por ejemplo, si agregamos una gota de tinta a un vaso con agua, la tinta difundirá a través del líquido y al cabo de un tiempo todo el vaso presentara una tinción pareja.
|
Fig. 4.17 - Difusión de una sustancia disuelta en un solvente. |
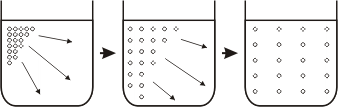
Para lograr esto no se requiere aporte externo de energía, sino que es suficiente con la energía cinética propia de las moléculas. Si tenemos en cuenta que la temperatura de un medio es, de alguna manera, un índice de la energía cinética de las moléculas presentes en el mismo, es fácil deducir que a mayor temperatura, más importante será el fenómeno de difusión.
Podemos definir entonces a la difusión como el movimiento de moléculas desde una zona de mayor concentración hacia una de menor concentración. A la diferencia de concentración que existe entre una zona y otra se la denomina gradiente.
a) DIFUSION SIMPLE
Cuando la difusión se realiza entre compartimientos separados por una membrana permeable a ese soluto, se denomina difusión simple y, como ya se dijo, no requiere de otra energía adicional que no sea el movimiento de las moléculas, desplazándose éstas a favor de su gradiente de concentración. En otras palabras, la difusión simple no requiere gasto de ATP, ya que es un fenómeno espontáneo. Las moléculas que se movilizan por difusión simple a través de la membrana son las no polares y pequeñas, las liposolubles y las polares pequeñas, pero sin carga eléctrica neta, como el H2O.
En el caso particular del H2O, la difusión simple se denomina ósmosis. El pasaje de agua a través de la membrana u ósmosis se lleva a cabo siempre en forma espontánea y muy rápidamente. El H2O difundirá desde el compartimiento de menor concentración de solutos o medio hipotónico, al de mayor concentración de solutos o medio hipertónico, de modo tal de igualar las concentraciones en ambos compartimientos. Al cabo de un tiempo, el resultado serán dos medios isotónicos, o sea, la concentración a ambos lados de la membrana será la misma.
|
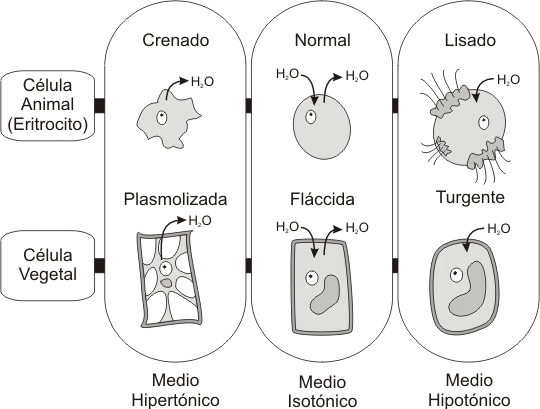
Fig. 4.18 -Efecto del proceso osmótico sobre una célula viva. |
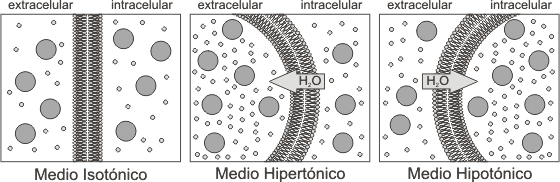
Si colocamos una célula, por ejemplo un glóbulo rojo, en una solución hipertónica (agua salada, por ejemplo) el H2O tenderá a salir por ósmosis hacia el medio extracelular, encogiendo o crenando al glóbulo rojo. En cambio, si el medio extracelular es hipotónico (agua destilada, por ejemplo) el H2O penetrará en la célula, hinchándola y, finalmente, ocasionando su ruptura o lisis. Cabe hacer aquí una breve aclaración: un medio no es por sí mismo ni hipertónico ni hipotónico; siempre que se use esta terminología lo que se esta haciendo es comparar un medio con respecto a otro. Por ejemplo, A puede ser hipertónico con respecto a B y, al mismo tiempo, A también puede ser hipotónico con respecto a C. Es decir, A tiene una concentración de solutos intermedia. Por otra parte, se dice que dos medios son isotónicos cuando su concentración de solutos es la misma.
Más adelante veremos (en Acuaporinas) que además de la ósmosis existen otros tipos de transporte de H2O a través de las membranas biológicas.
|
Fig. 4.19 - Osmosis. Efecto de los cambios de concentración de soluto en (a) células animales y (b) células vegetales |
b) DIFUSIÓN FACILITADA
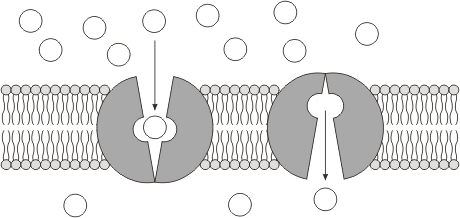
Aquellas moléculas que no pueden atravesar fácilmente las membranas por difusión simple debido a su polaridad y/o a su tamaño (por ej. glucosa, aminoácidos, iones, etc.), podrán hacerlo si están presentes sus respectivos transportadores. Dichos transportadores son proteínas integrales de membrana y se los puede agrupar del siguiente modo:
· Proteínas canal o canales iónicos
· Proteínas carrier o permeasas
La difusión facilitada ocurre siempre a favor del gradiente, por lo tanto no requiere gasto de energía adicional. Sin embargo, puede tratarse de un gradiente de concentración (las moléculas se dirigen del compartimiento de mayor concentración hacia el de menor concentración) o de un gradiente de potencial eléctrico (el soluto con carga eléctrica, independientemente de su signo, se desplazará de una zona donde la carga sea mayor hacia otra donde la carga sea menor).
Estas proteínas transportadoras presentes en las membranas presentan características muy similares a las enzimas:
· Saturabilidad (se saturan al alcanzar la máxima velocidad de transporte)
· Especificidad (reconocen a sus ligandos a través de un sitio específico)
· Pueden ser inhibidas por determinadas sustancias.
Cuando las proteínas transportadoras se saturan de solutos a transportar, alcanzan su máxima velocidad de transporte y por lo tanto las moléculas a ser transportadas deberán esperar a que se desocupen los sitios de unión.
b1) Canales iónicos:
Los canales iónicos son poros o túneles formados por una o varias proteínas transmembrana. En general, son de tipo multipaso, con un interior hidrofilico. Existen canales iónicos en todas las células, tanto en la membrana plasmática como en las membranas de los organoides. Son altamente selectivos, porque cada canal sólo puede transportar un tipo de ion (K+, Na+, etc.). Los iones se mueven a través del canal a una velocidad muy elevada (108 iones por segundo).
El transporte de un ion es impulsado por el gradiente electroquímico. O sea que un ion puede difundir de un lado a otro de la membrana, gracias a la diferencia de concentración como a la diferencia de carga eléctrica a ambos lados de la membrana.
La mayoría de los canales no permanecen abiertos permanentemente, sino que se abren en respuesta a estímulos. Estos estímulos pueden ser tanto la presencia de una sustancia inductora como una modificación de la carga eléctrica de la membrana (modificación del potencial eléctrico). Los canales que se abren o cierran en presencia de sustancias inductoras (ligandos) son llamados dependientes de ligando y los otros, dependientes de voltaje.
|
Fig. 4.20 - Diferentes tipos de canales. |

b2) Carriers o permeasas:
Al igual que los canales iónicos, las permeasas están formadas por proteínas transmembrana multipaso. Suelen transportar una gran variedad de iones como el HCO3- y otras moléculas polares sin carga como la glucosa.
Este tipo de proteínas fijan una única molécula de sustrato (o unas pocas) a la vez, y a continuación sufren un cambio conformacional reversible que les permite transportar el soluto de un lado al otro de la membrana (translocación). Aquí vale hacer otra aclaración: para entender la difusión facilitada no hay que pensar si una sustancia entra o sale de la célula, lo importante es considerar que se está movilizando algo a favor del gradiente (químico o eléctrico) gracias a la acción de proteínas transportadoras. Por esta razón es que no se requiere de energía adicional, no se requiere gasto de ATP, ya que es el propio gradiente el que impulsa el pasaje a través de los transportadores.
Este tipo de transporte es siempre sin gasto de energía y a favor del gradiente electroquímico. La velocidad de transporte es muy inferior al de los canales iónicos.
|
Fig. 4.21 -Transporte facilitado por medio de una permeasa. |
Existen tres tipos de permeasas:
-MONOTRANSPORTADORA O UNIPORTE: Transfieren UN solo tipo de soluto de un lado al otro de la membrana. (ej.: transporte de glucosa en la mayoría de las células animales, desde el medio extracelular, la sangre, donde la concentración es mayor, hacia el interior de las mismas donde es menor)
-COTRANSPORTADORA O SIMPORTE: Transfieren DOS tipos de solutos, ambos en el mismo sentido.
-CONTRATRANSPORTADORA O ANTIPORTE: Transfiere DOS tipos distintos de solutos en sentidos contrarios. Es decir, uno ingresa al citoplasma si, y solo si, simultáneamente el otro sale.
|
Fig. 4.22 - Tres tipos de transporte mediados por proteínas transportadoras |
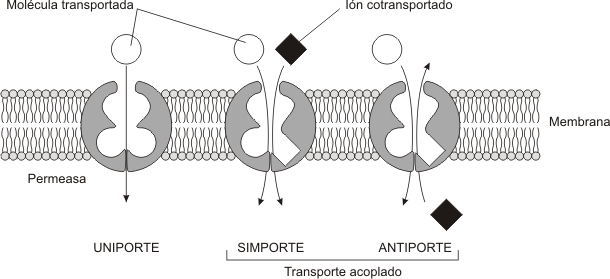
Los uniportes transportan las moléculas a favor de su gradiente de concentración. Como ejemplo podemos citar la glucosa y distintos aminoácidos. En cambio, los otros dos tipos de transporte acoplan el movimiento de un tipo de ion o molécula a favor de su gradiente de concentración con el de otro tipo de molécula o ion en contra de su gradiente de concentración. O sea lo que hacen es acoplar un transporte energéticamente favorable con otro que no lo es. Un ejemplo de COTRANSPORTE sería el transporte de Na+ y glucosa en la membrana plasmática de las células intestinales (ver más adelante) y uno de CONTRATRANSPORTE, el transporte de Cl- y HCO3- en la membrana de los glóbulos rojos.
Tanto el cotransporte como el contratransporte, son también llamados transportes acoplados, ya que no se pueden llevar a cabo si no están presentes ambos tipos de solutos.
Casos particulares de transporte pasivo: Ionóforos y Aquaporinas
IONOFOROS
|
Fig. 4.23 - Mecanismo de pasaje de iones a través de ionóforos transportadores móviles |
Estas sustancias tienen la propiedad de poder incorporarse a las membranas y aumentar la permeabilidad a ciertos iones. En general son fabricados por bacterias como mecanismos defensivos. Existen dos tipos distintos:
-Transportadores móviles: Se unen reversiblemente a un ion que se encuentra en el medio con mayor concentración, giran en la bicapa y lo liberan en el otro lado de la membrana. Ejemplo: Valinomicina (Fig.4.23).
- Formadores de canales: Son proteínas con estructura helicoidal, en cuyo interior de la hélice hay una región hidrofílica que permite el paso de iones monovalentes (con una sola carga eléctrica). Ejemplo: Gramidicina (Fig. 4.24).
|
Fig. 4.24 - Esquema de la estructura del canal de gramicidina |
ACUAPORINAS
Son canales especiales con estructura helicoidal que permiten el paso selectivo de H20. No son canales iónicos. En ciertas clases de células, por ejemplo en algunas células renales, se requiere un mayor transporte de H20 que el logrado exclusivamente con la difusión simple (osmosis). La estructura de las acuaporinas es semejante a la de los ionoforos formadores de canales.
c) TRANSPORTE ACTIVO
Las células no pueden depender únicamente del transporte pasivo dado que deben importar, por un lado, moléculas que están en menor concentración en medio extracelular que en el citoplasma y, por otro, necesitan mantener constante la composición iónica intracelular. Ambas funciones se llevan a cabo por medio del transporte activo.
Es un transporte que se realiza en contra del gradiente, ya sea este de concentración o eléctrico y, en consecuencia, se requerirá gasto de energía en forma de ATP.
El transporte activo se realiza por medio bombas y también presenta formas de monotransporte, cotransporte y contratransporte.
Posee las mismas características de especificidad y saturabilidad que la difusión facilitada, aunque difiere de ésta por realizarse contra el gradiente electroquímico. El transporte activo esta desfavorecido termodinámicamente (es endergónico) y se da solamente cuando está acoplado (directa o indirectamente) a un proceso exergónico como, por ej., la conversión de ATP a ADP + Pi. Debido a esto, las bombas se suelen denominar ATPasas de transporte.
Existen muchos tipos de ATPasas distintas. Aquí vamos a hablar de las más importantes, que son la Bomba de Na+-K+ (bomba sodio potasio)y la de K+/H+.
|
Fig. 4.25 - Esquema de la ATPasa. |
Las sustancias que se movilizan por transporte activo son en muchos casos las mismas que lo hacen a través de difusión facilitada, la diferencia fundamental es que en el primer caso lo hacen en contra del gradiente mientras que en el segundo lo hacen a favor.
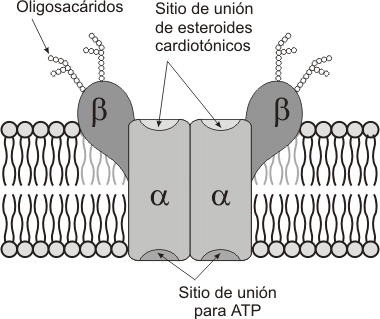
Bomba Na+/K+
Está presente en todas las membranas plasmáticas de las células animales. También se la conoce como Na+-K+ ATPasa. Es un complejo proteico formado por cuatro subunidades, todas ellas proteínas integrales de la membrana plasmática.
Su función es expulsar Na+ al espacio extracelular e introducir K+ al citosol. Ambos son movilizados en contra de su gradiente electroquímico, estableciendo así diferencias de concentración y carga entre el espacio extra e intracelular para ambos iones. Debido a que se esta transportando simultáneamente dos solutos distintos en sentidos opuestos, estamos en presencia de un sistema de contratransporte. Es importante recordar que, si bien el Na+ sale y el K+ ingresa a la célula, ambos lo hacen en contra de su gradiente y, en consecuencia, hace falta hidrolizar ATP para movilizarlos.
La Bomba Na+-K+ tiene simultáneamente funciones de proteína transportadora y de ATPasa (hidroliza ATP para obtener energía). Por lo menos un tercio de la energía que consume una célula animal se destina para impulsar esta bomba. En las células nerviosas, donde la actividad eléctrica es sumamente importante, este valor asciende al 60%. Cada ATPasa puede hidrolizar hasta 100 moléculas de ATP
Mecanismo de acción de la Bomba Na+/K
1) Tres iones de Na+ se unen al dominio citoplasmático de la ATPasa, debido a la gran afinidad que existe entre ambos.
2) Luego se hidroliza el ATP y se fosforila la proteína. Esto lleva a un cambio conformacional en la misma.
3) Esto permite la translocación de los iones Na+ hacia el espacio extracelular.
4) A continuación, dos iones K+ del medio extracelular, donde su concentración es menor, se unen a un sitio receptor de K+ accesible ahora desde el exterior de la célula. La unión del K+ con la proteína induce la liberación del fosfato.
5) La desfosforilación de la bomba, restituye la conformación original.
6) Esto permite la translocación de los iones K+ hacia el citoplasma. Se puede comenzar nuevamente el proceso. Por cada molécula de ATP que se hidroliza se posibilita el transporte de 3 iones Na+ hacia espacio extracelular y de 2 iones K+ al citoplasma.
Las transferencias de iones se hallan acopladas, y por lo tanto no pueden realizarse una independientemente de la otra.
|
Fig. 4.26 Mecanismo de acción de la ATPasa Na+/K+ |
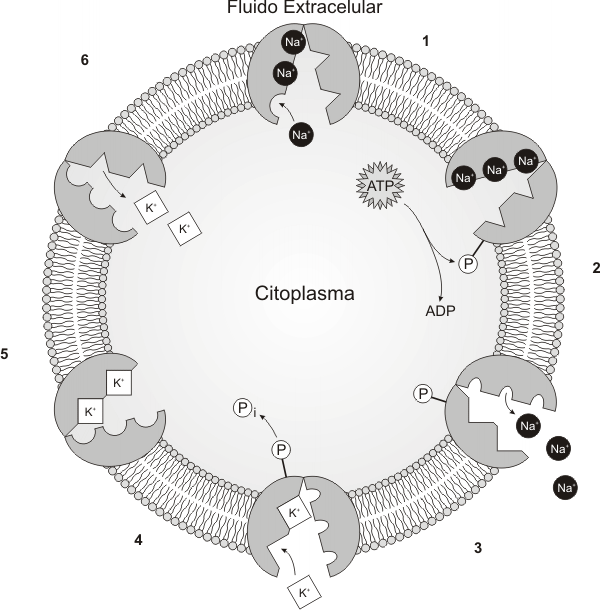
Las funciones de la bomba de Na+/K+ son:
a) Mantener diferencias en las concentraciones de Na+ y K+ intra y extracelulares.
b) Generar un potencial eléctrico de membrana, que es una diferencia de voltaje, o sea de carga, entre ambos lados de la membrana. Al bombear tres iones en una dirección y sólo dos en otra, se genera un potencial eléctrico negativo del lado interno de la membrana con respecto al externo. El lado citosólico es normalmente más negativo que el espacio extracelular.
c) Intervenir en la regulación del volumen celular.
d) Generar diferencias de concentración de Na+ o K+ para que otros transportadores pasivos utilicen indirectamente la energía potencial acumulada en este gradiente. Como ejemplo podemos citar al:
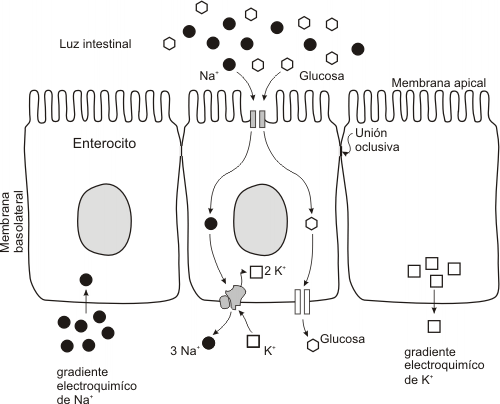
-COTRANSPORTE Na+/GLUCOSA (ya citado en difusión facilitada). Esta situación se da en las membranas apicales de las células del intestino delgado o en membranas de células renales, donde deberá absorberse glucosa desde la luz del intestino o de los túbulos renales, aunque las concentraciones extracelulares sean bajas. Gracias a la acción de la bomba Na+-K+ se expulsan iones Na+ a través de la membrana basal de la célula. De este modo, la concentración de Na+ intracelular se mantenida baja. En la región apical de la membrana se encuentra una permeasa pasiva cotransportadora de Na+ y glucosa. El Na+ ingresa de este modo a favor de su gradiente electroquímico al interior de la célula y arrastra a la glucosa con él, que ingresa de este modo en contra de su gradiente de concentración, gracias al sistema de cotransporte. Este tipo de transporte también se denomina transporte acoplado a gradientes iónicos o TRANSPORTE ACTIVO SECUNDARIO (ya que indirectamente está ligado a una bomba).
Posteriormente, la glucosa atravesará la célula y saldrá por difusión facilitada, a favor de su gradiente de concentración, hacia el torrente sanguíneo.
|
Fig. 4.27 Mecanismo de co-transporte Na+/glucosa en epitelio intestinal |
Hay otro tipo de ATPasa, presente en las membranas internas mitocondriales y de cloroplastos, que juega un papel muy importante en la obtención de la energía. Actúa como una ATPsintetasa (sintetiza ATP), gracias al gradiente de H+ que se genera a ambos lados de las membranas internas de cloroplastos y mitocondrias. Dicho proceso se verá con detenimiento en los capítulos de Respiración y fotosíntesis.
Existen otro tipo de bombas, como las de las membranas del Retículo endoplasmático liso de las células musculares, que se encargan de bombear iones Ca++ hacia el interior del REL y mantener baja la concentración citosólica Ca++ , o las de los lisosomas, que bombean H+ hacia el interior de los mismos, disminuyendo así el pH intralisosomal.
Hasta aquí analizamos el modo en el que los iones y las pequeñas moléculas atraviesan la membrana celular. Pero como ingresan o abandonan la célula partículas de mayor tamaño. Esto se realiza por medio del TRANSPORTE EN MASA. Este tipo de transporte involucra siempre gasto de ATP, ya que la célula realiza un movimiento general de su estructura (en particular de la membrana plasmática y del citoesqueleto -ver funciones del citoesqueleto-).
El mecanismo por medio del cual los materiales entran a la célula se denomina endocitosis y aquel por el cual la abandonan, exocitosis.
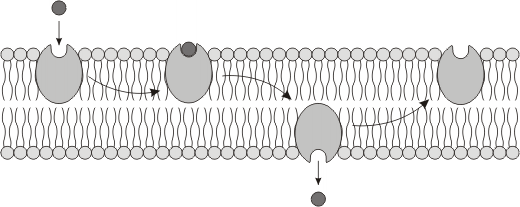
d1) ENDOCITOCIS
En este proceso una extensión de la membrana rodea progresivamente al material que será internalizado, luego se produce una gemación o invaginación de la membrana, y finalmente ésta se separa de la membrana, formando una vesícula endocítica. Posteriormente, el material incorporado es digerido por los lisosomas.
Las fibras de actina y miosina del citoesqueleto intervienen en este proceso.
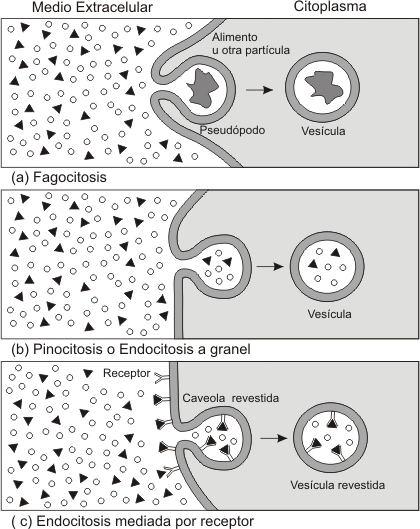
Se distinguen 3 tipos de endocitosis:
-Fagocitosis
-Pinocitosis
-Endocitosis mediada por receptor
A) Fagocitosis: Implica la ingestión de partículas de gran tamaño, como microorganismos, restos celulares, inclusive de otras células, por medio de vesículas llamadas fagosomas. Estos fagosomas suelen presentar un gran tamaño.
La fagocitosis sólo se da en determinados tipos de células. En algunos organismos unicelulares (protistas) constituye un modo de alimentación: engloban grandes partículas, por ej. bacterias, por medio de prolongaciones de la membrana plasmática llamados pseudópodos y las internalizan, formándose así un fagosoma o vesícula fagocítica. Posteriormente será degradada por las enzimas lisosomales. Para ampliar consultar en la bibliografía: Lisosomas.
En los animales sólo se da en algunas células altamente especializadas, llamadas células fagocíticas (macrófagos de los tejidos y glóbulos blancos sanguíneos denominados neutrófilos). En estos casos la función no es de índole nutricional, sino defensiva. Las células fagocíticas defienden nuestro organismo contra infecciones, ingiriendo microorganismos patógenos. Otra función sería eliminar células muertas o dañadas, o restos celulares (por ejemplo glóbulos rojos no funcionales). El proceso fagocítico se desencadena por la unión del material a endocitar con ciertos receptores de la membrana plasmática que reconocen al mismo.
B) Pinocitosis: Es la incorporación de fluído y de partículas disueltas en él por medio de pequeñas vesículas. Es un proceso inespecífico y la velocidad de ingestión es muy elevada. Por ejemplo, un macrófago puede ingerir por hora un cuarto de su volumen celular. El tamaño de estas vesículas endocíticas en mucho menor que el de los fagosomas.
C) Endocitosis mediada por receptor: En muchos aspectos es similar a la anterior, salvo que en este proceso, la endocitosis es mucho más selectiva. Determinadas moléculas (ligandos) que la célula desea incorporar son reconocidos por receptores específicos, ubicados en la membrana plasmática. Los ligandos se unen a estos receptores y estos complejos ligando-receptor confluyen, gracias a la fluidez de la membrana, a determinadas zonas de la misma, donde serán endocitados. La invaginación de la membrana se denomina en este caso fosita revestida. Esto se debe a que las vesículas presentan en su cara citosolica un revestimiento de proteínas características, en este caso de clatrina. La función de la misma, sería entre otras, permitir que se produzca la invaginación.
A continuación se forma la vesícula recubierta o revestida que se fusionará con un conjunto de vesículas llamadas endosomas, donde se clasifican las moléculas endocitadas y se las separa de los receptores.
Este proceso puede incrementar mil veces la eficiencia de internalización de un determinado ligando, sin tener que incrementar la absorción de fluido extracelular.
Un ejemplo importante de este proceso es la captación de colesterol por las células animales. El colesterol, debido a su carácter hidrofóbico, es transportado por la sangre unido a proteínas, formando complejos llamados lipoproteínas de baja densidad (LDL). Estas LDL se unen a receptores ubicados en la superficie celular y los complejos LDL-receptor son internalizados en vesículas revestidas y luego transferidas a los endosomas, previa liberación de la cubierta de clatrina. En el interior de los endosomas, el LDL se disocia del receptor y este es reciclado nuevamente a la membrana plasmática para captar nuevamente LDL.
|
Fig.4.28- Tipos de transporte en masa |
d2) EXOCITOSIS
Es el proceso inverso a la endocitosis. En este caso, material contenido en vesículas intracelulares también llamadas vesículas de secreción es vertido al medio extracelular.
La secreción de sustancias comienza generalmente con estímulos provenientes del medio extracelular, que inducen a las vesículas de secreción, ubicadas en las cercanías de la membrana, a fusionarse con la misma y volcar su contenido al medio extracelular. Así por ejemplo se liberan las proteínas de exportación (ver funciones del Aparato de Golgi) y los neurotransmisores (para ampliar esto ultimo consultar Sinapsis nerviosa en la bibliografía)
En este caso, la membrana de la vesícula pasa a formar parte de la membrana plasmática. Es decir, hay ganancia de membrana, mientras que en la endocitosis hay pérdida de membrana.
AUTOEVALUACIÓN
1. Esquematice el modelo de mosaico fluido de la membrana plasmática. Indique sus componentes.
2. Enumere las funciones más importantes de la membrana plasmática.
3. Conteste las siguientes preguntas con respecto a la estructura de la membrana plasmática:
a) ¿Cuál es la característica común a todos los lípidos de membrana?
b) ¿Por que se dice que la membrana plasmática es asimétrica?
c) ¿De qué depende el grado de fluidez de la membrana?
d) ¿Dónde espera encontrar más proteínas en la membrana interna de una mitocondria o en el retículo liso? ¿Por qué?
4. Resuelva el siguiente problema: En sus estudios sobre las células, Ud. descubre una nueva proteína, a la que llama esgfun. Esta proteína tiene un dominio A extracelular y un dominio C intracelular. Ud. descubre que esgfun es móvil, recorre lo largo de toda la membrana en 15 minutos. No importa cuantas veces la observe, el dominio A siempre está del lado extracelular y el C del lado intracelular.
a) Explique por qué se mantienen los dominios de esta manera.
b) Si Ud. fuera capaz de extraer todo el colesterol de esta membrana, qué cambios observaría en los movimientos de esgfingolípidos?.
5. Clasifique a los distintos tipos de transporte considerando los siguientes criterios :
a) Gasto de energía: pasivos (sin gasto) o activos (con gasto).
b) Uso de proteínas transportadoras: mediado (uso) o no mediado (no uso).
c) Número y dirección de partículas transportadas: uniporte, simporte y contratransporte.
6. Realice un cuadro comparativo donde indique las semejanzas y diferencias entre el transporte activo y la difusión facilitada.
7. Explique los distintos tipos de ionóforos.
8. Resuelva el siguiente problema: A Ud. se le provee de un cultivo de células en su medio de cultivo. Se encuentran en dicho medio muchas células por ml. Se le agrega una sustancia X y se puede medir la concentración interna de esta sustancia a través del tiempo de esta manera ud puede conocer como es tomada por las células. Describa (use gráficos recuerde los de enzimas) como podría determinar si X entró a la célula por difusión simple, transporte facilitado o transporte activo. Puede asumir que tiene al alcance de su mano todas las técnicas que necesita.
9. Conteste las siguientes preguntas sobre tipos de transporte
a) ¿Qué tipo de mecanismo utiliza la glucosa para ingresar a las células epiteliales del intestino desde la luz de este al interior de las mismas. ¿De dónde se obtiene la energía?
b) Los iones de Ca++ son eficientemente incorporados en muchas células vivientes, incluso cuando esto se realiza en contra de gradiente. Proponga un mecanismo para explicar esto.
c) Las neuronas y otras células excitables tienen membranas que son polarizadas. Existe una diferencia de voltaje que es negativo en el interior de la célula y positivo en el exterior. Explique cómo esta polarización es mantenida en una neurona en reposo. ¿Cuáles son los iones más importantes que participan? ¿existen iones más importantes que otros? ¿Cómo se crea y se mantiene esta diferencia de potencial?
d) Basado en sus conocimientos sobre los distintos tipos de transporte através de la membrana, proponga un mecanismo para explicar como estransportada la galactosa al interior de las células epiteliales del intestino.Incluya un diagrama de su mecanismo elegido (existe más de unaposibilidad, Ud. necesita solamente explicar uno)
e) ¿Cuáles son los distintos mecanismos por los que puede ingresar el agua y los iones en la célula?
f) Describa los mecanismos de transporte en masa y cite ejemplos.
Responda las siguientes preguntas de opción múltiple:
1. En que se diferencian las membranas de una célula eucariótica:
a- los fosfolípidos se encuentran solo en algunos tipos de membrana.
b- solo algunas membranas tienen permeabilidad selectiva.
c- solo algunas membranas tienen lípidos anfipáticos.
d- algunas proteínas son propias de cada membrana.
e- todas son correctas.
2. ¿Cuál de los siguientes procesos incluye todos los demás de la lista?
a- ósmosis.
b- difusión de un soluto a través de la membrana.
c- difusión facilitada.
d- transporte pasivo.
e- transporte de un ion a favor de gradiente.
3. Si una ameba es isotónica respecto a una solución que es hipertónica para un cangrejo, ¿en cuál de estos organismos ocurrirá un ingreso netos de agua al sumergir ambos en la solución?
a- en la ameba.
b- en el cangrejo.
c- en ninguno de los dos.
4. ¿Cuál de los siguientes factores podrían influir en la fluidez de la membrana?
a- una proporción grande de fosfolípidos insaturados.
b- una baja temperatura.
c- una proporción grande de fosfolípidos saturados.
d- un potencial alto de membrana.
e- ninguna es correcta.
5. Las células incorporan lipoproteínas utilizando:
a- fagocitosis
b- endocitosis mediada por receptor.
c- exocitosis.
d- transporte activo.
e- endocitosis a granel.
BIBLIOGRAFIA
- Alberts, B et al. (2002). Molecular Biology of the Cell. 4th edition.
- Brock, T. (1997) Biology of the Microorganisms. 8th edition. Prentice Hall. N.Y.
- De Robertis, E.; Hib, J.; (2001). Fundamentos de Biología Celular y Molecular. 3º Edición. El Ateneo. Bs.As.
- Harper,H. (1995). Manual de Bioquímica. Ediciones El Manual Moderno.
- Karp, G. (1998). Biología Celular y Molecular. Ed. Mc Graw Hill- Interamericana. México.
- Lehninger, A; Nelson, D; (1995). Principios de Bioquímica. 2º Edición. Ed. Omega. Barcelona.
- Lodish, H.; (2001). Biología Molecular de la Célula. 4º Edición. Ed. Panamericana. Bs.As.
- Maynard Smith, J; Szathmary, E. (2001). Ocho Hitos de la Evolución. Tusquets. Barcelona.
- Stryer, L. (2002), Biochemestry. 5th Ed. WH Freeman . NY
- Smith and Wood.(1998). Biología Molecular. Ed. Addisson-Wesley. Iberoamericana S.