Teorías de la Evolución y el Origen de las Especies
Viviana Sabatino - Andrea Lassalle- Silvia Márquez
¿QUÉ ES LA EVOLUCIÓN?
El término evolución significa desarrollo o desenvolvimiento, la transformación gradual de un estado a otro. El principio de la evolución biológica, que no es otra cosa que la aplicación de este concepto a los seres vivos, establece que los múltiples animales y vegetales que existen en la actualidad, descienden de organismos más simples, merced a modificaciones graduales que se han acumulado en el transcurso de las sucesivas generaciones. Las especies surgen de otras preexistentes.
PRIMERA SECCIÓN: LAS TEORÍAS DE LA EVOLUCIÓN¿QUÉ ES UNA TEORÍA DE LA EVOLUCIÓN?
Una teoría de la evolución es un conjunto de hipótesis y leyes que intentan explicar cómo pudo haberse producido ese cambio en el tiempo; ese conjunto debe estar coherentemente armado y no entrar en conflicto con otras áreas de la Biología.
La Biología contemporánea ubica a la teoría de la evolución corrientemente aceptada, la Teoría Sintética, como uno de sus pilares conceptuales más importantes. Es la única teoría general que, de una manera u otra, abarca prácticamente todos los campos de la Biología. Su peso está más desplazado hacia lo explicativo que hacia lo predictivo, dado que abarca aquéllo que la Biología tiene de histórico. Debe tenerse en cuenta que la Biología evolucionista no es primariamente una ciencia experimental. Es un punto de vista histórico con respecto a ciertos datos científicos. Es también un sistema de ideas del cual todo dato biológico es potencialmente integrante.
1. ANTECEDENTES DE LA TEORÍA SINTÉTICA
Actualmente ningún biólogo pone en duda el hecho de la evolución, pero no siempre ha sido así. En realidad, hace apenas doscientos años que se propuso por primera vez la idea de una historia de la evolución que comprende a todos los seres vivos del planeta. El primer intento científico y coherente de explicar su mecanismo, es decir cómo opera la evolución, se debe al naturalista francés Jean B. De Lamarck.
1.1. LA TEORÍA DE LAMARCK
La teoría de Lamarck, propuesta en 1809, se basa en las siguientes premisas:
1. Los organismos son guiados a través de su existencia por una fuerza innata y misteriosa que les permite sobreponerse a la adversidad del medio.
2. El medio ambiente actúa como una "fuerza modificadora" sobre los organismos, imponiendo necesidades que hacen surgir nuevos órganos y funciones.
3. El "uso y desuso de las partes": el desarrollo de los órganos y su actividad están en relación constante con el uso que de ellos se haga. Si un órgano es utilizado, entonces crece y se hace más eficiente. De lo contrario, puede degenerar y atrofiarse.
4. La herencia de los caracteres adquiridos: lo que se ha adquirido, impreso o modificado en la organización de los individuos durante el curso de su vida, es conservado y trasmitido a sus descendientes.
De acuerdo con este esquema la evolución podría ser más o menos como sigue. Supóngase que un determinado animal ancestral con el cuello corto se alimentaba con hojas de árbol. A medida que consumía las hojas inferiores del árbol estiraba el cuello para alcanzar las de más arriba. Debido a este estiramiento continuado a lo largo de toda la vida, el cuello se volvió algo más largo y entonces los descendientes heredaban un cuello algo más largo. Éstos a su vez se alimentaban de las hojas de los árboles y seguían estirando sus cuellos; y esto seguiría durante muchas generaciones. Con el tiempo llegó a formarse un animal con el cuello muy largo, semejante a una jirafa actual.
Esta teoría (en algunos aspectos incontrastable) tuvo gran éxito y contribuyó a la expansión de la idea de la evolución. Pero a la larga resultó insostenible. Es correcto que el uso y desuso conduce a la adquisición de caracteres. Por ejemplo, es de todos conocido que el ejercicio desarrolla músculos poderosos. Sin embargo Lamarck se equivocó al suponer que estas variaciones adquiridas (no genéticas) son heredables.
Los caracteres adquiridos no son heredables ya que son efectos producidos por el ambiente y el desarrollo, no por los genes. Sólo son heredables los caracteres genéticos y sólo si estos caracteres están regulados por genes presentes en células reproductoras. Lo que ocurra a otras células que no sean las gametas, a causa del uso y desuso, o por otras causas, no afecta a los genes de las gametas. Por consiguiente, a pesar de que Lamarck observó los efectos del uso y desuso correctamente en algunos casos, estos efectos no pueden tener un papel en la evolución.
1.2. LA TEORÍA DE DARWIN
El año en que Lamarck publicó su teoría -1809- fue también el año en que nació Charles Darwin. Durante su juventud, Darwin emprendió un viaje de cinco años de duración alrededor del mundo, como naturalista en la expedición del barco H. M. S. Beagle. Durante ella, hizo innumerables observaciones y recogió un gran número de plantas y animales distintos en muchas partes del mundo. Luego pasó casi veinte años examinando y estudiando los datos obtenidos. En el curso de este trabajo encontró pruebas para ciertas generalizaciones. Otro naturalista, Alfred Rusell Wallace, llegó básicamente a las mismas conclusiones independientemente, las cuales comunicó a Darwin.
En 1858 Darwin y Wallace juntos enunciaron una nueva teoría que sustituyó a la de Lamarck. Darwin también elaboró la nueva teoría en forma de libro. Este famoso trabajo, titulado "Sobre el origen de las especies mediante selección natural o la conservación de las razas favorecidas en la lucha por la vida" fue publicado en 1859.
En esencia, la teoría de Darwin se basa en algunas observaciones, y en conclusiones extraídas a partir de ellas.
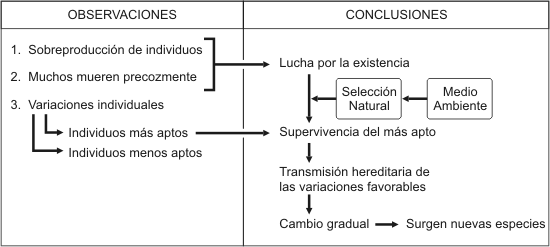
Darwin sabía que la superproducción de descendientes es la regla general en la naturaleza y que sólo unos pocos suelen sobrevivir. Entonces se pregunta: ¿En qué se diferencian los supervivientes de los que perecen? ¿Tienen características distintas, o es el azar el que determina los que sobreviven y los que mueren precozmente? ¿Es puro azar o se trata de un proceso dirigido cuyas causas pueden ser determinadas? Muchos son presa de sus enemigos, otros no consiguen suficiente alimento, no encuentran cobijo o, sencillamente, son desbancados por sus propios congéneres. Pero ¿y los que sobreviven? ¿Acaso son más veloces, están más camuflados quel os demás, son menos vulnerables a las enfermedades, aprovechan mejor su alimento, o tienen mejor olfato y pueden detectar al enemigo, o a sus presas, antes que otros miembros de su misma especie? Cuando la numerosa descendencia se enfrenta a las condiciones del medio ambiente, generalmente serán los "mejores", los "más aptos" los que logren sobrevivir. En esto consiste precisamente la " lucha por la existencia" -como había sido denominada ya por Malthus- denominación que fue adoptada por Darwin y que siempre se prestó a malentendidos. La situación queda mejor descrita por la expresión "supervivencia de los más aptos", de los mejor adaptados al entorno.
Así pues, Darwin observa que en la naturaleza se produce una selección análoga al proceso de la selección artificial que efectúa el criador de animales; la selección de los más aptos, de los más capacitados, que está determinada por las condiciones ambientales: es decir que existe una selección natural. Pero para que pueda tener lugar tal selección es necesario que se dé una superproducción de seres vivos.
Naturalmente, la selección sólo podrá darse si existen diferencias en las características de los descendientes de una misma especie; pero esto es algo que se da frecuentemente en todas las especies. Unos individuos resultan ser ligeramente más grandes, otros más pequeños; unos presentan una pigmentación más clara y otros más oscura; algunos pueden correr más rápidamente que el promedio de sus congéneres; algunas plantas presentan más ramificaciones que otras; este animal es más miedoso, aquél más agresivo... Estas diferencias, que Darwin denomina variaciones, pueden poner al ser vivo en situación ventajosa o desfavorable. En el último caso no estará en condiciones de superar las dificultades que le presente su entorno o de afrontar la competencia de otros seres vivos, por lo que pasará a formar parte de aquellos que sucumban antes de tiempo.
En cambio, las variantes favorables que mejoran la adaptación al medio de un ser vivo, permiten su supervivencia, su elección en el proceso de selección natural.
Los individuos que sobreviven dan origen a la próxima generación y de este modo las variaciones útiles se trasmiten a dicha generación y a las subsiguientes. Con el transcurso de muchos años de selección natural, los descendientes posteriores pueden resultar bien distintos de sus antecesores; esta diferencia puede ser suficientemente pronunciada como para que se los considere especies separadas.
Así, la selección natural se constituye en el principal agente del cambio evolutivo.
Podemos resumir lo hasta aquí expuesto de la siguiente forma:
Si aplicamos la teoría a un ejemplo: La teoría de Darwin sobre la selección natural de las especies se basa en cuatro nociones fundamentales. Comprenderemos más fácilmente su doctrina si aplicamos esas cuatro ideas a una población viva.
1- El número excesivo de las crías
Todas las especies son capaces de producir un número de crías superior al que bastaría para ser sustituidas.

Una pareja de ratones puede dar seis camadas por año, de seis crías, más o menos, cada una. A las seis semanas de nacidas, éstas pueden a su vez reproducirse.

Imaginemos cuántos ratones habría si todos ellos sobrevivieran y siguieran reproduciéndose.

¿Por qué la Tierra no está llena de ratones? Aunque una pareja de ratones puede producir muchas más crías de las que se necesitan para sustituirla, el número de miembros de cada población tiende a seguir siendo más o menos el mismo, debido a que no todas las crías sobreviven el tiempo necesario para reproducirse.
2- La lucha por la supervivencia
El medio ambiente puede influir en las probabilidades individuales de supervivencia.
Entre todos los seres vivos y el medio ambiente existe una relación recíproca. El entorno proporciona alimentos, espacio y un territorio adecuados para la vida, pero alberga también animales rivales o depredadores. De ahí que no haya población alguna cuyos individuos sobrevivan en su totalidad hasta poder reproducirse.

Un ratón puede ser devorado por un animal.

O puede resultar incapaz de encontrar alimento suficiente.

O puede no encontrar pareja.
3- Diferencias individuales importantes
Dado que no todos los individuos son idénticos, algunos tienen más probabilidades de sobrevivir que otros.

No hay dos ratones exactamente iguales y algunas de las variaciones que existen entre ellos pueden influir en las probabilidades que tienen para sobrevivir.
No todos los ratones son del mismo color: unos son más oscuros que otros.

Sobre un fondo oscuro, es más fácil distinguir a los ratones pálidos, de modo que es más probable que éstos sean devorados por los búhos. Los ratones más oscuros están mejor adaptados al medio y tienen mayores probabilidades de sobrevivir.
4- Una cuestión de herencia
Algunas características se transmiten a la generación siguiente.

En una superficie oscura del suelo es menos probable que los animales de presa distingan a los ratones de piel oscura. Así, sus características son las que más probablemente se transmitirán a la generación siguiente.
En la próxima generación habrá una proporción mayor de ratones de piel oscura que en las anteriores.
Si se mantienen las mismas condiciones, continuará aumentando la proporción de ratones de color oscuro dentro de la población.

Después de muchas generaciones es probable que aumente la proporción de individuos bien adaptados. Darwin llamó a este proceso la SELECCIÓN NATURAL.
La selección constituye la explicación del fenómeno en virtud del cual las características de una población pueden modificarse a medida que los individuos se adaptan mejor a su medio ambiente
Muchas personas tienen a menudo la impresión de que esta teoría es la explicación moderna de la evolución. Esto no es exacto. Más aún, se repiten, como hechos ciertos, ideas erróneas que tuvieron su origen en una mala interpretación o tergiversación del darwinismo, tales como "el hombre desciende del mono", "la supervivencia del más fuerte" o "la ley de la jungla".
De lo postulado por Darwin, sigue vigente la idea de la selección natural, aunque ya se ha puesto en claro que tiene poco que ver con una "lucha".
Además, la selección natural es sólo parte del mecanismo evolutivo, ya que al igual que Lamarck, Darwin fracasó en la identificación de las causas de la variación. Darwin comprendió que la conservación de las variaciones dentro de la población era esencial para su teoría de la selección natural, pero nunca supo cómo se conservaba esta variación. Igual que muchos otros científicos de su época, pensaba que si los dos progenitores de un individuo diferían entre sí en algún carácter, los gametos que este individuo produjera serían uniformes y transmitirían el carácter en alguna condición intermedia. Este tipo de herencia eliminaría muy rápidamente las diferencias entre individuos. Su efecto sería análogo al de mezclar y remezclar agua y tinta en muchos recipientes hasta que todos contuvieran una solución coloreada algo diluida. Este punto de vista sobre la herencia es erróneo. Los dos alelos que lleva un heterocigota se segregan en gametas sin contaminación mutua. Los genes que un individuo transmite a su prole pueden estar en combinaciones nuevas, pero son los mismos que recibió de sus progenitores. Si Darwin hubiese conocido los experimentos de Mendel, se habría podido dar cuenta de que su problema de explicar la persistencia de la variación dentro de las poblaciones no existía.
En resumen, la explicación de Darwin (y Wallace) fue incompleta. Pero, aun en su limitación, fue la primera en indicar la dirección correcta.
ORIGEN Y EVOLUCIÓN DE LAS JIRAFAS SEGÚN LAS IDEAS DE LAMARCK (izquierda) Y DARWIN (derecha)
|
Las jirafas primitivas de cuello corto y patas de longitud normal, intentaban constantemente -sobre todo en épocas de sequía- ramonear las verdes hojas de los árboles. A causa del continuo estiramiento del cuello y las patas estos miembros comenzaron a alargarse. 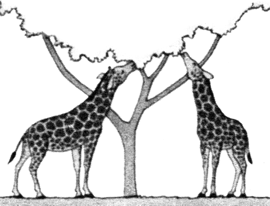 |
Entre las antiguas jirafas se daban algunas diferencias en cuanto a la longitud de sus cuellos y patas; estas variaciones se transmiten hereditariamente. 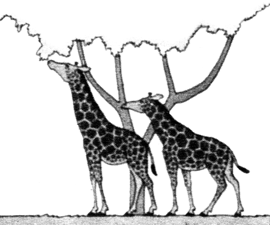 |
|
Sus descendientes empezaron a nacer con el cuello y las patas más largos, y el ejercicio y "afán" de estirarse hizo que sus miembros continuaran estirándose cada vez más. 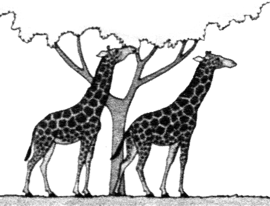 |
Las jirafas más altas de cada generación disfrutaban de la ventaja de alcanzar más alimentos que las pequeñas, por lo que se reproducían mucho más intensamente. 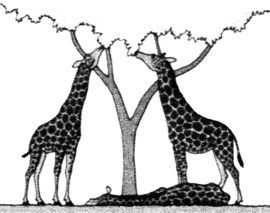 |
|
Debido a este continuo estiramiento, llegaron con el tiempo a convertirse en las jirafas que conocemos hoy día con sus cuellos y patas largos. 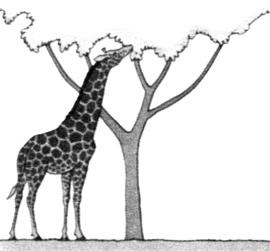 |
Con el tiempo la selección natural favorece a los ejemplares de cuellos y patas largos. 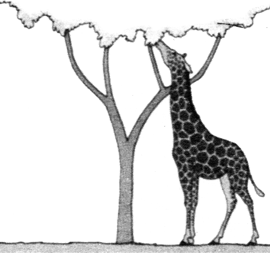 |
2. LA TEORÍA SINTÉTICA
Aunque algunos científicos del siglo XIX aceptaron de inmediato los argumentos de Darwin sobre la selección natural como principal agente del cambio evolutivo, otros muchos se adhirieron a teorías distintas, entre ellas la lamarckiana.
Luego, cuando se redescubrió la genética mendeliana, a comienzos del siglo XX, surgió una nueva escuela: la de los mutacionistas. Éstos veían a la evolución como si procediera de súbitos estallidos de cambio, desatados por acontecimientos internos, a saber: las mutaciones. Los efectos del ambiente, mediados por la selección, quedaron relegados –en el mejor de los casos- a un papel secundario.
En las primeras décadas de este siglo la Biología Evolutiva se hallaba sumida en cierta confusión intelectual. Sin embargo, en los años treinta y cuarenta, comenzó a aflorar un consenso que luego sería conocido como Síntesis Moderna, términos acuñados por Julian Huxley, nieto de Thomas Henry Huxley, amigo y tenaz defensor de Darwin.
La Síntesis Moderna es el producto de la unión entre el campo de la Genética, en rápida maduración, y las ideas más fundamentales del seleccionismo, todo ello en el contexto de la Genética de poblaciones.
La Teoría Sintética llevó a una rara unanimidad a esta rama de la ciencia, tradicionalmente tumultuosa.
Los principales artífices de la teoría Sintética fueron, junto con Huxley, Theodosius Dobzhansky, Ernest Mayr, Sewell Wright, Ledyard Stebbins, George Gaylord Simpson y Bernard Rensch.
2.1. EL MECANISMO EVOLUTIVO SEGÚN LA TEORÍA SINTÉTICA
Los cambios en el curso de la evolución ocurren sobre los organismos individuales. Sin embargo, sólo son observables en el seno de los grupos y en el transcurso de sucesivas generaciones. Por esto, los cambios que importan a la evolución son aquéllos heredables y la unidad de evolución no es el individuo sino la población. Se entiende por tal a una asociación de individuos de la misma especie que coexisten en un hábitat y, lo más importante, que pueden cruzarse entre sí, son potencialmente interfértiles. Dada esta potencial interfertilidad o panmixia (del griego pan: todo y mixis: mezcla) los genes de los individuos que conforman una población fluyen libremente entre sus integrantes mediante el proceso de reproducción sexual.
La sumatoria de los genes de los individuos que componen una población es el patrimonio génico (llamado asimismo pool, complejo, reservorio o acervo génico) de la misma. El pool génico es una abstracción: obviamente, los genes de un pool se hallan en diferentes combinaciones de alelos, constituyendo genotipos de individuos particulares. Las Leyes de Mendel nos explican la forma en que estos alelos se transmiten en la reproducción sexual, determinando el genotipo de cada individuo.
Siendo que el fenotipo –es decir las características morfofisiológicas y etológicas de un organismo- está determinado en gran medida por su genotipo, la herencia se comporta hasta cierto punto como un factor conservador, que tiende a mantener las generaciones sucesivas dentro de un esquema común.
Pero en poblaciones naturales, el pool génico se ve enriquecido en cada generación por nuevas variantes génicas. Esta variación genética es el producto de la mutación, que introduce nuevos alelos y de la reproducción sexual, que favorece la recombinación de los genes existentes, creando diversos genotipos. Así, la existencia de variación genotípica (sumada a factores exógenos) acarrea la variación fenotípica.
Como señalara Darwin, entre las “variaciones individuales” algunos fenotipos resultan más ventajosos que otros frente a determinadas condiciones ambientales. Éstos serán los “elegidos”, los favorecidos por la selección natural. En efecto, es el medio quien da la medida del grado de adaptación de cada fenotipo. Pero, ¿qué se entiende por aptitud en términos evolutivos? La aptitud no es sólo la capacidad de confrontar exitosamente con las dificultades del ambiente, sino gracias a ello y además de ello, alcanzar la posibilidad de reproducirse. Esto significa que la generación siguiente heredará los alelos determinantes del fenotipo seleccionado. En consecuencia, dichos alelos –en tanto las condiciones del medio no varíen- irán aumentando su frecuencia en el acervo génico de la población.
Según la Teoría Sintética, evolución es, precisamente, el cambio progresivo en la frecuencia de alelos de un patrimonio genético a lo largo del tiempo.
· La Teoría Sintética colocó a la selección natural en un papel central, como una fuerza evolutiva que modela a los organismos, permitiendo una creciente adaptación al medio generación tras generación. Si bien la selección natural elige fenotipos, lo que se transmite a la descendencia es la base genética de aquéllos. Por lo tanto, la materia prima sobre la cual trabaja la selección es la variación genética de la población. Por esto mismo, se dice que la evolución es oportunista; no persigue como fin la adaptación perfecta, pues trabaja sobre la materia prima disponible en la población. Y cuanto mayor sea la variabilidad genética de una población, más materia prima se ofrecerá a la evolución.
· En su versión más extrema, la Síntesis considera que los seres son capaces de una variabilidad genética (y por consiguiente fenotípica) infinita, por lo que la selección modela a una especie a partir de un conjunto continuo e infinito de posibilidades. En consecuencia, el resultado de la “todopoderosa” selección sería la adaptación casi perfecta de los organismos a su ambiente.
· La Síntesis rescata la idea de la evolución como resultado de la acumulación gradual de pequeños cambios adicionales.
Cabe destacar que en el seno de la Teoría Sintética, en continua revisión, algunas de estas hipótesis – el papel siempre protagónico de la selección natural, la capacidad infinita de variabilidad genética, así como el carácter gradual del cambio evolutivo- han sido cuestionados por diferentes corrientes surgidas con posterioridad.
2.2. LAS FUENTES DE VARIABILIDAD GENÉTICA
2.2.1 MUTACIÓN
Definida en sentido estricto, la mutación es la alteración al azar de la información que llevan los genes. La mutación puede afectar los genes de diversas maneras y en una extensión variable. En cualquiera de los casos, resulta el único mecanismo generador de información heredable nueva que se conoce. (Aunque si reducimos nuestro ámbito a una sola población, también puede aparecer información nueva por la inmigración o ingreso de individuos provenientes de otras poblaciones).
La frecuencia de mutación puede ser modificada por diversas influencias, tales como la presencia de ciertos otros genes, o la aplicación de radiaciones o de sustancias químicas, pero la naturaleza de la nueva información es aleatoria, no está determinada por estas influencias.
Esa información nueva no es necesariamente positiva para un individuo, también puede resultar perjudicial o neutra. No obstante, la aparición de mutaciones favorables, aunque se produzcan con baja frecuencia, alcanza para que la variación heredable siga teniendo “combustible” (en los seres humanos se calcula que cada recién nacido es portador de dos mutaciones nuevas).
De acuerdo con el grado de extensión que alcancen las mutaciones, éstas pueden ser:
a. Mutaciones génicas: afectan a la estructura de un gen, por sustitución, deleción o inserción de bases en uno o más tripletes.
b. Mutaciones cromosómicas: afectan a la estructura de un cromosoma, por pérdida o separación de una de sus partes que, a veces, puede volverse a unir al mismo o a otro cromosoma, con alteraciones en mayor o menor grado.
c. Mutaciones genómicas o cariotípicas: en las que si bien los cromosomas individuales permanecen inalterados, el cambio afecta a la dotación cromosómica en conjunto, ya que el número de cromosomas aumenta o disminuye.
2.1.2 RECOMBINACIÓN
La selección natural sólo discrimina fenotipos y éstos son el resultado de la acción cooperativa de todos los genes que constituyen el genotipo. Para que un alelo sea seleccionado, no basta con que sea “bueno” en sí mismo; es necesario además que establezca una armónica combinación con sus “compañeros de viaje”. Si gen muy ventajoso compartiera el genotipo con un alelo letal, también sería eliminado; por el contrario, un gen neutro podría propagarse por el sólo hecho de hallarse ligado a otro favorecido por la selección.
Richard Dawkins, en su libro “El gen egoísta”, establece una analogía entre los genes y un grupo de remeros. Un remero que se vale sólo de sus propios medios no puede ganar una regata. Remar un bote es una empresa cooperativa. El éxito dependerá de qué tan buenos sean los otros miembros del equipo y de la coordinación que exista entre ellos. Siguiendo la analogía de Dawkins, la recombinación equivale a formar equipos de remeros, escogiendo sus integrantes al azar de entre un grupo de ellos (el pool génico). Cabe la posibilidad de que algunos equipos resulten muy malos, pero también es posible que en algún momento surja un equipo que reúna a los mejores.
En las poblaciones, las recombinaciones, ocurridas al azar, son el resultado de la reproducción sexual y tienen lugar en tres instancias:
a. Recombinación gamética: en una población panmíctica, cualquier individuo en edad reproductiva puede cruzarse con otro del sexo opuesto. Esto quiere decir que si en la población hay dos mil individuos en estas condiciones, existen, potencialmente, un millón de cruzamientos (combinaciones) diferentes.
b. Recombinación cromosómica: como se vio en el estudio de la meiosis, la contribución de cromosomas paternos o maternos al complemento haploide de las gametas es un fenómeno aleatorio. Un individuo produce gametas que llevan informaciones sensiblemente diferentes entre sí.
c. Recombinación génica: es consecuencia del crossing-over.
La reproducción sexual, por los mecanismos expuestos, permite la creación de innumerables genotipos nuevos, ofreciendo a las especies el beneficio de una rápida adaptación. En esto reside su ventaja sobre la reproducción asexual, que transmite la combinación genética sin ninguna alteración.
En ciertos casos, sin embargo, la panmixia se ve restringida. Por ejemplo, en las poblaciones pequeñas es frecuente el cruzamiento entre parientes y en las plantas es muy común la autofecundación. Ambas son formas de incesto y limitan la recombinación. El efecto del incesto en una población es la tendencia a la homocigosis (predominio de individuos homocigotas). De esta manera, alelos recesivos que pudieran haberse mantenido ocultos se manifiestan fenotípicamente. Si dichos alelos reducen la posibilidad de adaptación de sus portadores, el incesto resulta nefasto. Pero si, por el contrario, la población ha logrado una combinación genética óptima para las condiciones de su hábitat, la conservación de la condición homocigótica es de vital importancia.
En las poblaciones heterogaméticas o panmícticas se mantiene un porcentaje alto de heterocigosis (predominio de heterocigotas). Esto implica una gran diversidad genética que se recombina en cada generación. Aunque muchas de estas recombinaciones no sean productos o genotipos óptimos, la capacidad de adaptación y la flexibilidad evolutiva alcanzan su máximo.
El genotipo heterocigota es, en muchos casos, más favorable que cualquiera de los homocigotas. A esta superioridad del heterocigota se la llama vigor híbrido o heterosis. No obstante, la población mantiene un polimorfismo balanceado de fenotipos, es decir, los homocigotas no son eliminados, ya que siempre surgen en un 50% de la cruza de dos heterocigotas.
La heterocigosis supone, además, una gran reserva de alelos recesivos ocultos que, por homocigosis, pueden emerger y ser presentados a la selección natural. Suele ocurrir que alelos surgidos por mutación y que permanecen ocultos y neutros por mucho tiempo en un pool génico, emerjan en un momento en que cambian las condiciones ambientales, pasando a ser ventajosos (recordemos el carácter “oportunista“ de la evolución). Este fenómeno se conoce como preadaptación.
2.3. SELECCIÓN NATURAL: REPRODUCCIÓN DIFERENCIAL
La selección organiza y ordena la materia prima suministrada por las mutaciones y recombinaciones y para ello se orienta según la “dirección” que marcan los cambios del ambiente. La “lucha por la existencia” es el instrumento que emplea. Pero esta lucha no se desarrolla de un modo cruento y brutal, se trata más bien de un proceso estadístico en el que algunos individuos dejan más descendientes que otros. Dicho proceso se denomina reproducción diferencial y en ello consiste la concepción actual de la selección natural.
La aptitud, entonces, está determinada por la capacidad de propagar los alelos propios en las generaciones siguientes, a expensas de otros alelos diferentes.
Según este punto de vista, el organismo mayor y más poderoso de una población podría ser muy “apto” en cuanto a comportamiento, pero si fuera estéril, carecería de importancia en el aspecto reproductor y, por lo tanto, en el evolutivo. Recíprocamente, un organismo débil, enfermizo, podría ser “inepto” en comportamiento, pero tener una descendencia numerosa. Claro que, generalmente (aunque no necesariamente) los que dejan más descendientes son los organismos mejor preparados para sobrevivir en su ambiente. Por lo tanto, la reproducción diferencial tiende a imponer en la población la información genética de los fenotipos más ventajosos.
2.4. UN EJEMPLO DE EVOLUCIÓN RECIENTE: EL DDT Y LAS MOSCAS
Para ilustrar la forma en que interactúan los distintos agentes que participan del proceso evolutivo, describiremos un caso de evolución reciente que fue observado por el hombre.
En 1948, el químico suizo Paul A. Muller recibió el premio Nobel por el descubrimiento del DDT. Este insecticida provocó euforia por su efectividad, aunque ésta fue efímera, pues pronto los insectos se hicieron inmunes a su acción.
¿Qué había ocurrido?
Se descubrió que entre los insectos libres se daban casos de mayor o menor resistencia al insecticida (variación fenotípica). Es decir que frente a la presión selectiva ejercida por el DDT, algunos individuos resultaban más aptos.
Por otro lado, experimentos efectuados con la mosca de la fruta demostraron que los rasgos que conferían resistencia eran heredables; estaban inscriptos en el genotipo y existía variación genética para ellos. Dichos rasgos se transmitían, en principio, en forma independiente. Ellos son:
1. Capa de quitina tres veces menos permeable al DDT que en otros insectos.
2. Presencia de enzimas que les permiten transformar al DDT, muy rápidamente, en un compuesto inocuo.
3. Mayor cantidad de grasa corporal donde se acumula el DDT sin que cause daño.
4. Mayor tamaño, por ende soportan dosis mayores.
5. Comportamientos tales como abandonar las casas a la noche (cuando habitualmente se fumiga) y evitar las superficies verticales (que son las más rociadas).
Los individuos que no presentaban ninguno de estos rasgos perecieron en las primeras fumigaciones: fueron eliminados por selección natural.
Los que tenían alguno de ellos, en cambio, sobrevivieron y se cruzaron entre sí para dar origen a la siguiente generación (reproducción diferencial), que heredó así los alelos para la resistencia. Éstos, que habían aparecido en el pool génico por mutación y que hasta el descubrimiento del DDT carecieron de importancia adaptativa, aumentaron su frecuencia progresivamente al ser favorecidos por la selección. Esto es un ejemplo de preadaptación.
Con el correr de las generaciones no sólo aumentaba el número de individuos resistentes, sino también el grado de resistencia. Dado que los sobrevivientes portaban al menos uno de los alelos ventajosos, al entrecruzarse generaban descendientes que reunían en su genotipo los genes heredados por ambas líneas (recombinación gamética). Las gametas de estos insectos podían llevar a la vez genes “buenos” paternos y maternos (recombinación cromosómica). Por último, sus nietos heredaban los alelos de ambos orígenes ligados en el mismo cromosoma, gracias al proceso de crossing-over (recombinación génica).
En este caso se puede leer claramente cómo a partir de un pool con alta variabilidad genética, la presión selectiva del medio y la reproducción sexual y diferencial generan cambios en las frecuencias de alelos y por consiguiente en las características fenotípicas de una población. En el marco de la Teoría Sintética esto es evolución.
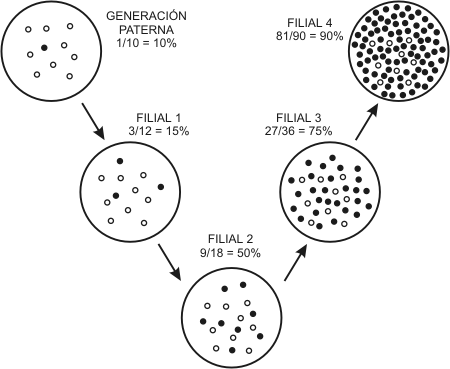
2.5. GENÉTICA DE POBLACIONES: LA LEY DE HARDY–WEINBERG
Hemos visto que la población es el centro de estudio de las investigaciones acerca de la evolución; la Genética de poblaciones es la disciplina que lleva a cabo tales investigaciones. Sus objetos de estudio son:
· Los cambios que se producen en el patrimonio genético de las poblaciones.
· El análisis de las causas que provocan dichos cambios.
Para encarar este estudio parte de una población ideal, cuyo comportamiento es descripto por el principio de Hardy–Weinberg. Éste fue enunciado en 1908, en forma simultánea e independiente por el matemático inglés Hardy y el médico alemán Weinberg. Se deduce de la aplicación de las leyes de Mendel a las poblaciones y predice lo que ocurrirá con sus frecuencias génicas bajo ciertas condiciones.
Lo analizaremos mediante un ejemplo, para luego enunciarlo:
Consideremos una población cuyos individuos presentan un rasgo regido por un par de alelos: A y a. Los genotipos posibles serán AA, aa y Aa. Dadas las siguientes proporciones en la población inicial:
N = Nº total de individuos = 100
|
Genotipos |
AA |
aa |
Aa |
|
N° de individuos |
36 |
16 |
48 |
¿Qué hemos de esperar para la siguiente generación, suponiendo que todos los individuos se cruzaran al azar?
Primero averiguararemos la frecuencia de los alelos A y a en las gametas.
Llamaremos: p a la frecuencia del alelo A
q a la frecuencia del alelo a
Como sólo existen dos alelos: p + q = 1
Dado que cada individuo AA aportará un gen A y cada individuo Aa aportará un gen A o un gen a, con igual probabilidad (50%), la frecuencia p se calcula con la siguiente fórmula:
p = AA + ½ Aa = 36 + ½ 48 = 0,6
N 100
Con la misma fórmula calculamos q:
q = aa + ½ Aa = 16 + ½ 48 = 0,4
N 100
Una vez halladas p y q, puede calcularse la proporción de los genotipos en la generación descendiente:
|
Óvulo |
Espermatozoide |
Cigoto |
Frecuencia |
Porcentaje |
|
|
A |
A |
AA |
p x p = 0,6 x 0.6 = 0,36 |
36 % |
|
|
a |
a |
aa |
q x q = 0,4 x 0,4 = 0,16 |
16 % |
|
|
A |
a |
Aa |
p x q = 0,6 x 0,4 = 0,24 |
24 % |
48% |
|
a |
A |
Aa |
p x q = 0,6 x 0,4 = 0,24 |
24 % |
|
De estos simples cálculos se desprende que la población obtenida es genéticamente igual a su predecesora inmediata; las frecuencias de los alelos en el pool no han variado.
De aquí surge el enunciado de Hardy–Weinberg:
“Si el apareamiento es aleatorio, si no ocurren mutaciones y si la población es grande, entonces las frecuencias génicas de una población permanecen constantes de generación en generación.”
Una población que cumpla con las condiciones de esta ley tendrá un ritmo de evolución igual a cero. Pero esto no ocurre con ninguna población natural; en ellas las frecuencias de alelos siempre están cambiando.
El valor de la ley consiste en proporcionar un patrón que nos permite detectar los cambios y las fuerzas responsables de los mismos. Cualquier desviación de las condiciones que establece la ley actuará como factor de evolución:
· El apareamiento NO es al azar en la mayoría de las poblaciones naturales. No todos los genotipos tienen el mismo valor adaptativo, por lo tanto algunos resultan más beneficiados que otros por la selección natural. Supongamos que el genotipo aa cause la muerte de un tercio de sus portadores en el período embrionario. Los cigotos aa se formarán pero no llegarán a la edad reproductora. Los individuos AA y Aa formarán una fracción mayor de la población reproductora: las frecuencias génicas se alteran. Por otro lado, las parejas se forman por criterios no aleatorios, tales como fuerza, inteligencia, apariencia, atractivo sexual. Esta selección sexual es una forma más de la selección natural y hará que ciertos genes se propaguen más que otros.
· Los genes mutan. La introducción de un alelo nuevo en la población produce automáticamente un cambio en las frecuencias preexistentes. Posteriormente, dicho alelo será sometido a la selección natural.
· El principio de Hardy–Weinberg no se cumple en poblaciones pequeñas (menos de 100 individuos). Esto es así porque el equilibrio de Hardy–Weinberg depende de las leyes de la probabilidad, que no se cumplen cuando el número de casos considerados es bajo, por lo que en pequeñas poblaciones pueden obtenerse frecuencias génicas diferentes de las esperadas.
2. 6. LA DERIVA GENÉTICA
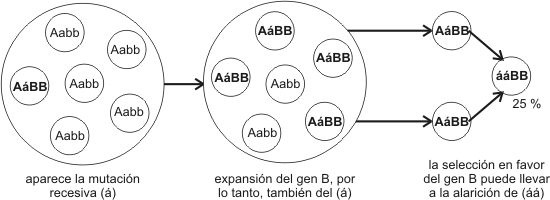
La deriva genética se define como la alteración al azar (independientemente del valor adaptativo) de la frecuencia de un gen en poblaciones pequeñas.
En la deriva genética se observan dos procesos:
§ En primer lugar, un grupo reducido de individuos queda separado de una población grande. La parte del pool genético que pasa a la nueva población puede o no ser representativa de la original (tal vez algunos alelos no estén presentes y otros se hallen representados en exceso).
§ En segundo lugar, el reducido tamaño de la población conduce al incesto, por lo que los alelos - que llegaron a ella por azar – tienden a recombinarse en forma homocigótica.
El papel de la deriva genética como agente evolutivo ha sido discutido, pero se conocen por lo menos dos situaciones en las que se demostró su influencia:
1. Efecto del fundador. Un ejemplo lo constituye el grupo Old Order Amish, fundado en Pennsylvania hace 200 años. En este grupo existe una altísima frecuencia de un alelo recesivo, que es poco frecuente en la población mundial y que en estado homocigótico ocasiona enanismo y polidactilia. El grupo se mantuvo aislado y desciende de unos pocos individuos, de modo que, por accidente, uno de ellos debió ser portador del alelo.
2. Cuello de botella. En algunas poblaciones el tamaño oscila a lo largo del año, pasando por períodos en que el número de individuos se mantiene en un nivel mínimo. Estas fluctuaciones ocurren espontáneamente en poblaciones de insectos y de mamíferos como los lemmings, aunque también pueden ser causadas por el hombre o por catástrofes naturales. Cuando la población queda reducida, se produce una deriva genética con la consiguiente homocigosis ocasionada por incesto, de forma que al aumentar la población masiva, el patrimonio génico resulta más o menos alterado. El tránsito por este cuello de botella, como todo fenómeno aleatorio, puede suponer un nuevo impulso hacia una mejor adaptación, o bien conducir a la población al fracaso.
SEGUNDA SECCIÓN: EL ORIGEN DE LAS ESPECIES1. LA ESPECIACIÓN
Los pequeños cambios que ocurren dentro de las poblaciones –la microevolución- pueden conducir al surgimiento de nuevas especies, fenómeno denominado especiación.
Las especies son grupos de poblaciones naturales que se cruzan o pueden entrecruzarse y se encuentran reproductivamente aisladas de otros grupos similares. El detalle esencial de esta definición es el aislamiento reproductivo. Sabemos que existen dos especies aparte cuando ambas pueden ocupar el mismo espacio sin entrecruzarse.
En términos de genética de poblaciones, los miembros de una especie comparten un reservorio génico netamente separado de los reservorios génicos de otras especies.
En la especiación, entonces, se presentan dos cuestiones importantes:
§ Una cuestión central es cómo un reservorio de genes se separa de otro para iniciar una trayectoria evolutiva aparte.
§ Otra cuestión subsidiaria es cómo dos especies, a menudo muy similares entre sí, habitan el mismo lugar al mismo tiempo, pero se mantienen reproductivamente aisladas.
Veamos cómo se resuelven ambas cuestiones en las dos formas de especiación conocidas: la especiación alopátrica y la especiación simpátrica.
1.1. ESPECIACIÓN ALOPÁTRICA
Este tipo de especiación se inicia con la separación de los individuos de una población a causa de la aparición de una barrera geográfica.
Las barreras geográficas son de muy distintos tipos. A menudo las islas se prestan para la aparición de especies nuevas; tal fue el caso de Australia cuando se fraccionó de Pangea. Otras barreras son menos obvias que los océanos. La aparición de áreas pantanosas entre dos hábitats puede ser una barrera efectiva; unos pocos metros de tierra seca pueden aislar a dos poblaciones de caracoles.
¿En qué forma el aislamiento geográfico conduce a la formación de nuevas especies?
Un aislamiento de este tipo tiene como consecuencia que el flujo genético se produzca separadamente en cada una de las poblaciones, dando lugar a un desarrollo independiente. Incluso el punto de partida para esta evolución será distinto: habrá genes del patrimonio genético original que estén más representados en un grupo que en el otro y viceversa. Además las mutaciones y combinaciones que ocurran estarán sometidas a diferentes presiones de selección, ya que los biotopos serán distintos. Primero surgirán dos razas distintas y tras un largo período de aislamiento llegarán a establecerse dos subespecies. Los cambios en el medio ambiente impulsarán a ambas subespecies a emigrar y colonizar nuevas áreas, e irán experimentando cada vez mayores divergencias genéticas, hasta llegar a constituir dos especies distintas que habitan en diferentes regiones, es decir dos especies alopátricas (del griego allos: diferente y del latín patria: hogar).
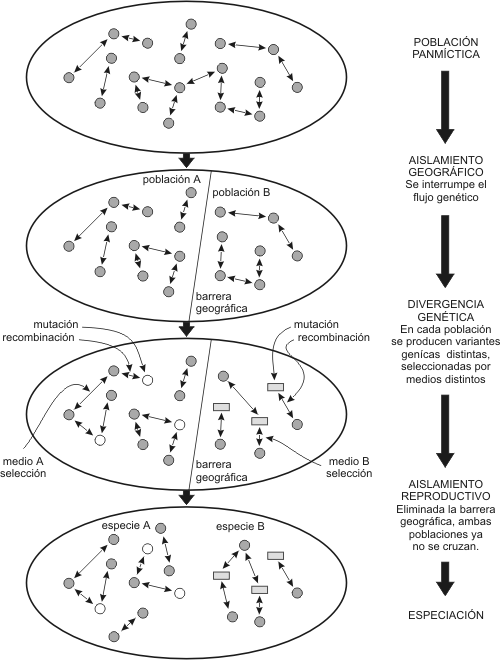
Fig. 15.3 – Especiación alopátrica
Si posteriormente se llegara
a producir el encuentro de ambas especies en un territorio común, ya no
sería posible el entrecruzamiento. Una vez que el aislamiento geográfico
ha favorecido la divergencia y la especiación, empiezan a actuar un sinnúmero
de otros mecanismos aisladores: son los mecanismos de aislamiento
reproductivo. No todos los mecanismos de aislamiento actúan en
todos los casos, pero sirven para fortalecer los límites entre las especies.
Cualquier combinación favorable alcanzada en una de las especies se “diluiría”
si el flujo genético se restableciera entre ellas.
Los mecanismos de aislamiento reproductivo son:
|
Precigótico |
ecológico |
Preapareamiento |
|
estacional |
||
|
etológico |
||
|
mecánico |
Posapareamiento |
|
|
gamético |
||
|
Poscigótico |
genético |
|
|
esterilidad del híbrido |
|
¿Cuáles son los factores que impiden el cruzamiento de dos poblaciones? Dos poblaciones son incapaces de cruzarse si . . . |
|||||||
|
|
 |
|
|||||
 |
 |
||||||
...viven en hábitats diferentes |
. . .tienen aspectos diferentes |
. . .tienen un comportamiento diferente |
|||||
 |
 |
 |
 |
 |
|||
 |
|||||||
|
. . . se reproducen en distintas estaciones del año |
. . sus gametas son incompatibles |
. . . su descendencia no sobrevive o es estéril |
|||||

Fig. 15,4 – Aislamiento reproductivo
Aislamiento ecológico. Si dos poblaciones son capaces de aparearse, pero no lo hacen porque viven en nichos ecológicos distintos, se dice que están aisladas ecológicamente. Por ejemplo: dos poblaciones de ciervos son capaces de entrecruzarse en el laboratorio. En la naturaleza una población vive del bosque y otra de un campo cercano. Cada población permanece casi completamente dentro de su propio nicho. En esta situación son prácticamente nulas las posibilidades de entrecruzamiento.
Aislamiento estacional. Aunque dos poblaciones habiten una misma región, sus períodos reproductores pueden ocurrir en distintas estaciones del año. Por ejemplo: en el norte de Europa, el saúco rojo florece dos meses antes que el saúco negro; sin embargo es posible cruzar las dos especies artificialmente.
Aislamiento etológico. Se debe a diferencias de comportamiento. Por ejemplo: en ciertos peces, aves e insectos, el macho y la hembra llevan a cabo rituales de cortejo muy elaborados, en los que cada movimiento de uno desencadena una respuesta específica del otro. Estos comportamientos vienen determinados genéticamente, y cualquier cambio en el ritual haría que el otro miembro de la pareja perdiera interés.
Aislamiento mecánico. Una pequeña modificación anatómica, aparentemente mínima, puede ser suficiente para evitar la cópula.
Aislamiento gamético. En animales se manifiesta mediante mecanismos modificadores de la acidez de las vías genitales, que ciertos tipos de gametas no pueden soportar, o a través de mecanismos inmunitarios anti-gametas extrañas.
Aislamiento genético. El hecho de que se produzca la fusión de las gametas no asegura que el cigoto pueda desarrollarse. Un cigoto producto de especies diferentes, en la mayoría de los casos no pasa de las primeras divisiones.
Esterilidad del híbrido. El caballo y el asno pueden cruzarse dando un híbrido, la mula. La prole sobrevive, es fuerte, pero no puede reproducirse.
1.1. ESPECIACIÓN SIMPÁTRICA
Es un tipo de especiación que puede ocurrir sin aislamiento geográfico (del griego sym: junto y del latín patria: hogar).
Los únicos mecanismos de especiación simpátrica conocidos hasta el momento consisten en una hibridación seguida de poliploidía ,o bien en una autopoliploidía , y se verifican en vegetales.
El genetista ruso Karpetschenko realizó el experimento de cruzar plantas de distintas especies: rábanos y coles. De estos cruzamientos obtuvo algunos híbridos en la f1, pero casi todos resultaron estériles. Sólo unos cuantos llegaron a producir semillas y de éstas se desarrollaron plantas que se comportaron como una nueva especie (con algunas características del rábano, otras de la col y otras intermedias, y que no se cruzaban con las especies progenitoras).
¿Cómo fue posible? Ambas especies paternas poseen 2n=18 cromosomas; cada una de sus gametas tiene por lo tanto n=9 cromosomas. Así pues, los híbridos f1 tendrán 2n=18 cromosomas: 9 proceden del rábano y 9 de la col. Estos cromosomas han de formar pares de homólogos y, puesto que no lo son, la meiosis se produce con dificultad. Sin embargo, hay algunos casos aislados en los que la meiosis de los híbridos f1 se desarrolla de modo diferente. Pudo observarse que en la formación de los granos de polen no se llegaba a producir la primera meiosis, que normalmente reduce el número de cromosomas a la mitad. Así, los 9 cromosomas del rábano y los 9 de la col permanecían juntos y en la segunda división meiótica daban lugar a dos granos de polen, cada uno con 18 cromosomas. Naturalmente, esto puede ocurrir también en la formación de óvulos. Los híbridos f2 tenían 36 cromosomas: 18 del rábano y 18 de la col. Por lo tanto eran tetraploides y precisamente por eso podían efectuar una meiosis normal, puesto que los 18 cromosomas de la col formaban ahora 9 parejas de cromosomas homólogos y lo mismo ocurría con los provenientes del rábano. La poliploidía es un ejemplo de mutación genómica.
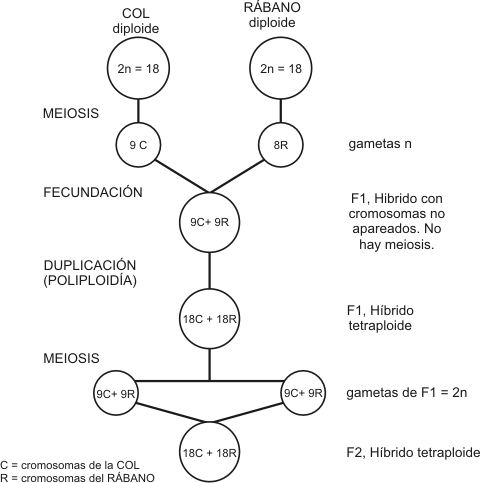
Fig. 15.5 – Especiación simpátrica. Híbrido poliploide de col y rábano (cruza de Karpetschenko)
Sin duda Karpetschenko había
producido una nueva especie y no hay nada de artificial en su experimento:
más o menos la mitad de las 235.000 especies de plantas con flor tuvieron
un origen poliploide y muchas especies agrícolas importantes, entre ellas
el trigo, son poliploides híbridos.
2. LA MACROEVOLUCIÓN
La macroevolución es una evolución por encima del nivel de especie, con emergencia de nuevos taxones superiores de organismos. Por ejemplo, el surgimiento del primer vertebrado mamífero, a partir de un antecesor que no lo era o, mucho antes, la aparición del primer insecto.
¿Cómo se explica la macroevolución? El único terreno probatorio de una teoría macroevolucionista es el registro fósil y toda conclusión sobre la evolución a gran escala debe corroborarse con este registro.
Diferentes interpretaciones de las pruebas fósiles han producido una de las controversias más importantes dentro de la Síntesis. El gradualismo y el equilibrio puntuado (o equilibrio interrumpido) representan dos modelos diferentes con respecto a este tema.
El gradualismo es el modelo macroevolucionista ortodoxo. Explica la macroevolución como el producto de un cambio lento, de la acumulación de muchos pequeños cambios en el transcurso del tiempo. Este cambio gradual debería reflejarse en el registro fósil con la aparición de numerosa formas de transición entre los grupos de organismos. Sin embargo, el registro no es abundante en formas intermedias. Los gradualistas atribuyen esta discrepancia entre su modelo y las pruebas halladas a la imperfección del propio registro geológico (según expresiones de Darwin, el registro geológico es una narración de la que se han perdido algunos volúmenes y muchas páginas).
El modelo del equilibrio puntuado – propuesto en 1972 por N. Eldredge y S.J. Gould- sostiene en cambio que el registro fósil es un fiel reflejo de lo que en realidad ocurrió. Las especies aparecen repentinamente en los estratos geológicos, se las encuentra en ellos por 5 a 10 millones de años sin grandes cambios morfológicos y luego desaparecen abruptamente del registro, sustituidas por otra especie emparentada, pero distinta. Eldredge y Gould utilizan los términos estasis e interrupción, respectivamente, para designar estos períodos. Según su modelo, las interrupciones abruptas en el registro fósil de una especie reflejarían el momento en que ésta fue reemplazada por una pequeña población periférica –en la cual el ritmo de evolución habría sido más rápido- que compitió con la especie originaria y terminó por sustituirla. De acuerdo con este patrón, la selección natural no sólo opera dentro de la población, sino también entre especies, y los cambios cualitativamente importantes en los organismos ocurrirían en períodos relativamente breves –desde el punto de vista geológico-separados por largos períodos de equilibrio.
LECTURA COMPLEMENTARIA
Ciencia al Día
LA EVOLUCIÓN HOY
La gran síntesis neodarwiniana, que en la primera mitad de este siglo logró conciliar la genética con la teoría de la evolución, giraba, hasta cierto punto, en el vacío. Si bien había quedado establecido que los genes eran las unidades y los portadores de la herencia y se los había logrado ubicar en los cromosomas presentes en el núcleo de cada célula, nadie sabía exactamente de qué estaba hecho un gen. En 1954 Watson y Crick encontraron, finalmente, la estructura de los genes: grandes y largas moléculas de ADN (ácido desoxirribonucleico) enrolladas en forma de doble hélice. Este descubrimiento es el hecho central de la biología en este siglo, y naturalmente abrió un nuevo capítulo que, con justicia, se llamó biología molecular, y entre muchísimas otras cosas, obligó a replantear –en niveles más profundos aún- la teoría sintética de la evolución, a la vez que lanzó grandes desafíos a la misma.
Porque si la Teoría Sintética postulaba modificaciones (mutaciones) que daban lugar a la aparición de un nuevo gen, que luego, selección mediante, se extendía por el patrimonio genético de la especie, la lupa provista por la biología molecular exigía ahora una explicación – y una explicación molecular- de por qué y cómo evolucionan y se modifican los propios genes. La palabra “mutación” mostró su enorme complejidad: al lado de las mutaciones puramente al azar, aparecieron otros mecanismos portadores del cambio y la evolución genética. Por ejemplo, la duplicación en tándem de partes de un gen, o la fusión de diversos genes para formar un gen único (y nuevo). O la extraña aparición de información genética cruzada: genes de una especie que aparecen en otra independiente y no conectada mediante una cadena evolutiva. (Aunque se sospecha que los agentes de esta transmisión informativa son pequeños trozos de ADN circular conocidos como plásmidos, capaces de acarrear material genético de una especie a otra, este fenómeno todavía permanece en la oscuridad.)
Toda esta avalancha de nuevos datos moleculares multiplicaron las discusiones y tendencias entre los evolucionistas. Algunos investigadores sostienen que, a nivel molecular, los cambios no se producen por el puro azar que postula la Teoría Sintética, sino que habría alguna especie de determinismo – en algún remoto sentido lamarckiano – capaz de guiar la evolución del ADN. En el otro extremo, la teoría neutralista, de Motoo Kimura del Instituto Nacional de Genética en el Japón, sostiene que es el puro azar el que gobierna la propagación de un gen nuevo o modificado en una población y que la selección natural no juega absolutamente ningún papel, lo cual implica un serio cuestionamiento a la Teoría Sintética. A su vez, entre quienes aceptan el lugar central del mecanismo de selección natural, las aguas se dividen apenas aparece la pregunta: ¿selección natural entre quiénes? Para el darwinismo tradicional y el neodarwinismo, entre individuos. Pero Stephen Jay Gould, Eldredge, entre otros partidarios del equilibrio puntuado, se inclinan por la selección natural entre especie (véase esta misma columna del 15/5/90). En el otro extremo, Richard Dawkins sostiene que la lucha por la existencia y la supervivencia del más apto se da de manera exclusiva entre genes y que toda envoltura, como las células, o las complicadas estructuras biológicas de nuestros cuerpos, no son sino artimañas de los genes para asegurar su permanencia: la gallina no es sino el medio que tiene un huevo para garantizar otro huevo, decía en el siglo pasado aquel exquisito satírico y darwiniano Samuel Butler.
Así, la Teoría de la Evolución está en un fecundo momento de confrontaciones y discusiones, de sugerencias, experimentos y réplicas. Esta posición sostiene que todas las objeciones pueden ser subsumidas en una Teoría Sintética levemente modificada: aquella, que las modificaciones deben ser radicales; la de más allá, que hay que revisar de manera completa todos los resultados de la genética de poblaciones. De una manera o de otra, todas las corrientes del evolucionismo actual se encaminan, a marcha forzada, hacia una nueva síntesis. Fiel a su nombre, la Teoría de la Evolución parece un objeto vivo, que cambia y se transforma. De la pionera historia lamarckiana a la genial concepción de Darwin, de Darwin a la Teoría Sintética, de la Teoría Sintética a las diversas posturas que generó la biología molecular, ha demostrado una ágil versatilidad, una profunda capacidad de adaptación y una serena belleza. El vigoroso mecanismo que transformó las primeras y tímidas moléculas capaces de duplicarse en el multiforme panorama de las especies, que incluyen seres que piensan viajar a las estrellas no se conoce todavía perfectamente. Pero nos acercamos cada vez más, y sentimos el estremecimiento que produce contemplar la vida reproduciéndose, perpetuándose y cambiando a través de los eones.
Leonardo Moledo
Fuente: Diario Clarín.
Publicada el 22-05-90
ACTIVIDADES DE AUTOEVALUACIÓN
1. La teoría de Darwin:
a) se basó en conocimientos avanzados de genética
b) se apoyó en las leyes de Mendel
c) destacó el papel de las mutaciones
d) introdujo el concepto de selección natural
e) negó la posibilidad de herencia de los caracteres adquiridos
2. “Los antecesores de los caballos actuales tenían un mayor número de dedos. El uso de las extremidades para la carrera en terrenos duros, condujo a la reducción del número de dedos del caballo actual”. Esta explicación corresponde a la teoría:
a) de Lamarck
b) de Darwin
c) sintética
d) saltacionista
e) neutralista
3. “Algunas bacterias poseían información genética que, al descubrirse los antibióticos, les resultó útil para resistir el efecto de éstos. Dichas bacterias aumentaron su frecuencia en las poblaciones.” Esta explicación corresponde a la teoría:
a) de Lamarck
b) de Darwin
c) sintética
d) saltacionista
e) neutralista
4. “Algunas moscas poseían información genética que, al descubrirse el insecticida DDT, les resultó útil para resistir el efecto de éste. Dichas moscas aumentaron su frecuencia en las poblaciones”. Esta explicación corresponde a la teoría:
a) de Lamarck
b) de Darwin
c) sintética
d) del equilibrio puntuado
e) neutralista
5. “El uso de las patas para la natación dio lugar a la aparición de membranas interdigitales en las ranas”. Esta explicación corresponde a la teoría:
a) de Lamarck
b) de Darwin
c) sintética
d) del equilibrio puntuado
e) neutralista
6. El cambio de frecuencias génicas dentro de un pool génico, a causa del azar, se denomina:
a) selección natural
b) deriva génica
c) aislamiento
d) hibridación
e) selección sexual
7. La sustitución de una base en un gen se denomina mutación...
a) cariotípica
b) cromosómica
c) génica
d) ninguna es correcta
8. La reproducción sexual produce variabilidad genética como consecuencia de:
a) recombinación gamética
b) recombinación cromosómica
c) recombinación génica
d) todas son correctas
9. Un individuo bien adaptado a su medio tiene mayores oportunidades de reproducirse que los demás. Es probable que sus genes se propaguen dentro de la población. Este hecho es un ejemplo de:
a) mutación
b) recombinación
c) selección natural
d) deriva génica
e) especiación
10. ¿Cuál de los siguientes mecanismos de aislamiento reproductivo es poscigótico?
a) estacional
b) ecológico
c) incompatibilidad gamética
d) incompatibilidad anatómica
e) esterilidad del híbrido
11. ¿Cuál de los siguientes mecanismos de aislamiento reproductivo es menos económico para una especie? Justifique su respuesta.
a) estacional
b) ecológico
c) incompatibilidad gamética
d) incompatibilidad anatómica
e) esterilidad del híbrido
12. La genética y el mecanismo de selección natural son los pilares en los que se asienta la teoría:
a) de Lamarck
b) de Darwin
c) sintética
d) del equilibrio puntuado
e) neutralista
13. La especiación alopátrica implica la secuencia:
a) aislamiento geográfico - divergencia genética - aislamiento reproductivo
b) aislamiento reproductivo - divergencia genética - aislamiento geográfico
c) ausencia de barreras - hibridación
d) hibridación - poliploidía
e) cuello de botella - aislamiento reproductivo
14. La especiación simpátrica implica la secuencia:
a) aislamiento geográfico - divergencia genética - aislamiento reproductivo
b) aislamiento reproductivo - divergencia genética - aislamiento geográfico
c) presencia de barreras - hibridación
d) hibridación - poliploidía
e) cuello de botella - aislamiento reproductivo
15. Con respecto a la macroevolución, la teoría sintética ortodoxa propone un mecanismo:
a) a saltos
b) gradual
c) de equilibrio puntuado
d) neutralista
e) todas son correctas
16. Los elefantes marinos fueron diezmados por la caza intensiva en el siglo pasado. A partir de 20 ejemplares sobrevivientes logró recuperarse la población, que hoy supera los 30.000 individuos. ¿En qué fenómeno, desde el punto de vista evolutivo, encuadraría esta situación? ¿Cómo piensa que pudo afectar la diversidad genética de la población? ¿Por qué?
17. Darwin observó que ciertas adaptaciones de los animales no están relacionadas con la supervivencia, sino con la posibilidad de aparearse. Tal es el caso del plumaje llamativo del pavo real macho, destinado a atraer a las hembras. Él llamó a este fenómeno selección sexual, para diferenciarlo de la selección natural. Explique por qué, en el marco de la teoría sintética, Ud. podría considerar a la selección sexual una forma más de la selección natural. ¿Se cumple la ley de Hardy-Weinberg en este caso?
BIBLIOGRAFÍA
1. Schwoerbel, Wolgfgang; 1984; Evolución; Biblioteca Científica Salvat; N° 43.
2. Curtis, Helena; 1985; Biología; Ed. Panamericana; 4° Edición.
3. Simpsom, G. G.; 1984; El sentido de la evolución; EUDEBA.
4. PROMEC; 1985; Biología; Estructura Modular 5, Módulo 2.
5. Gould, S. J.; 1986; El pulgar del panda; Biblioteca de Divulgación Científica Muy Interesante; N° 16.
6. Dawkins, R.; 1985; El gen egoísta; Biblioteca Científica Salvat; N° 9.
7. Lewin, R.; 1987; Evolución humana; Biblioteca Científica Salvat; N° 64.
8. Stansfield, W.; 1992; Genética; Mc Graw-Hill; 3° Ed.
9. Ayala,F.; 1994; La naturaleza inacabada, Ensayos en torno a la evolución; Biblioteca Científica Salvat.
10. Weisz, P. B.; 1975; La Ciencia de la Biología; Ed. Omega.
11. El Correo de la Unesco; Mayo de 1982; De ratones y mariposas nocturnas.
12. British Museum Natural History; 1981; El origen de las especies; Ed.Akal.