EL NÚCLEO CELULAR
Silvia Márquez- Andrea Lassalle- Viviana Sabbatino- Gladys Gálvez
IntroducciÓn
El núcleo es la estructura más destacada de la célula eucarionte, tanto por su morfología como por sus funciones. Su tamaño es variable (5 a 10 mm) al igual que su ubicación siendo en la mayoría de los tipos celulares central.
El núcleo tiene tres funciones primarias, todas ellas relacionadas con su contenido de ADN. Ellas son:
1. Almacenar la información genética en el ADN.
2. Recuperar la información almacenada en el ADN en la forma de ARN.
3. Ejecutar, dirigir y regular las actividades citoplasmáticas, a través del producto de la expresión de los genes: las proteínas.
En el núcleo se localizan los procesos a través de lo cuales se llevan a cabo dichas funciones. Estos procesos son:
1. La duplicación del ADN y su ensamblado con proteínas (histonas) para formar la cromatina.
2. La transcripción de los genes a ARN y el procesamiento de éstos a sus formas maduras, muchas de las cuales son transportadas al citoplasma para su traducción y
3. La regulación de la expresión genética.
Estructura del núcleo (Fig. 10.1)
El núcleo está rodeado por la envoltura nuclear, una doble membrana interrumpida por numerosos poros nucleares. Los poros actúan como una compuerta selectiva a través de la cual ciertas proteínas ingresan desde el citoplasma, como también permiten la salida de los distintos ARN y sus proteínas asociadas.
La envoltura nuclear es sostenida desde el exterior por una red de filamentos intermedios dependientes del citoesqueleto, mientras que la lámina nuclear, la cual se localiza adyacente a la superficie interna de la envoltura nuclear, provee soporte interno.
El núcleo también tiene un nucleoplasma, en el cual están disueltos sus solutos y un esqueleto filamentoso, la matriz nuclear la cual provee soporte a los cromosomas y a los grandes complejos proteicos que intervienen en la replicación y transcripción del ADN.
Los cromosomas aparecen ocupando lugares específicos. Los genes que codifican productos relacionados, aunque estén localizados en diferentes cromosomas, pueden estar ubicados próximos en el núcleo interfásico. Por ejemplo, los cromosomas humanos 13, 14, 15, 21 y 22 poseen un gran número de genes que codifican para ARNr. Dichos cromosomas están agrupados de tal forma que los genes de los ARNr están todos juntos y confinados en el nucléolo, el lugar donde se sintetizan, procesan y ensamblan los ARNr. Esta separación física asegura que los ARNr puedan ser eficientemente ensamblados dentro de las subunidades ribosomales.
En el núcleo, los genes transcripcionalmente activos tienden a estar separados de los inactivos. Los activos se encuentran ubicados centralmente, mientras que los silentes están confinados próximos a la envoltura nuclear.
Tan pronto como las células entran en mitosis o meiosis, los fragmentos de la matriz nuclear dirigen la condensación de los cromosomas, constituyéndose en la parte central de los mismos.

Fig. 10.1 - Esquema de un núcleo interfásico
La envoltura nuclear
La envoltura está formada por dos membranas concéntricas interrumpidas por poros nucleares y por la lámina nuclear.
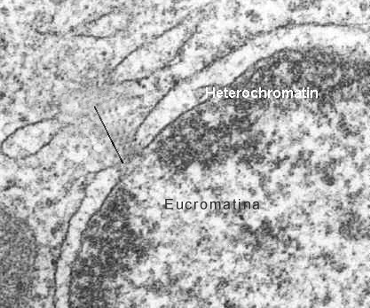
Fig. 10.2- Microfotografía electrónica de la envoltura nuclear
Las membranas delimitan un espacio de 10 a 50 nm, el espacio o cisterna perinuclear. La membrana externa en contacto con el citoplasma tiene ribosomas adheridos, que sintetizan las proteínas que se vuelcan al espacio perinuclear. El espacio perinuclear se continua con el REG.
La membrana interna posee proteínas integrales que le son propias, que se unen a la lámina nuclear y a los cromosomas.
La lámina nuclear, capa fibrosa de 10 a 15 nm en la que apoya la membrana interna, está formada por proteínas del tipo de los filamentos intermedios, polímeros de lamina o laminina nuclear. Ellas se unen a las proteínas integrales de membrana.
La fosforilación de las laminas provoca el desensamble de la lámina nuclear causando la ruptura de la envoltura al inicio de la división celular.
La lámina nuclear confiere estabilidad mecánica a la envoltura nuclear. Además, al interactuar con la cromatina participa en la determinación de la organización tridimensional del núcleo interfásico.
Si bien la formación de la lámina no se requiere durante los pasos iniciales, la organización de la envoltura es indispensable para el crecimiento posterior y el mantenimiento de su integridad. Las laminas se incorporan luego que la cisterna perinuclear rodea al ADN y se inicia el transporte entre el núcleo y el citoplasma. (Fig. 10.3)
La envoltura nuclear es un derivado del sistema de endomembranas, siendo esto evidente al inicio de la división celular, cuando la envoltura se desorganiza y pasa a formar parte del sistema de cisternas y vesículas del retículo endoplásmico.
La aparición de la envoltura nuclear permitió que los eucariontes aislaran los procesos genéticos principales, como la autoduplicación del ADN o la síntesis de ARN. Además esto posibilitó que el ARNm se modifique dentro del núcleo antes de ser traducido en los ribosomas. Estas modificaciones no ocurren en los procariontes, ya que a medida que la ARN polimerasa sintetiza el ARN, simultáneamente el extremo 5’ se une al ribosoma y comienza la traducción.
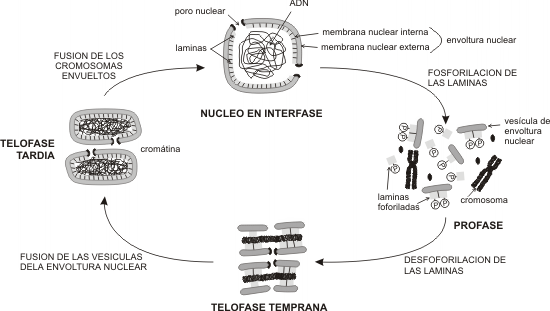
Fig. 10.3 - Mecanismo de formación y desintegración de la membrana nuclear
Complejos de poro nuclear
La envoltura nuclear presenta estructuras discoidales llamadas complejos de poro nuclear (CPN) (Fig. 10.4).
El número de CPN es variable, incrementándose a medida que aumenta la actividad celular. En una célula de mamífero hay entre 3000 a 4000 complejos de poro. Cada CPN es una estructura macromolecular compleja constituida por un gran número de proteínas de disposición octamérica. Está formado por:
· Ocho columnas proteicas, que forman las paredes laterales del poro.
· Un anillo externo, formado por ocho unidades proteicas.
· Un anillo interno, también con estructura octamérica.
· Proteínas de anclaje que fijan cada columna al espacio perinuclear.
· Proteínas radiales que se proyectan desde las columnas hacia la luz del poro, a manera de diafragma
· Proteínas fibrilares fijas al anillo interno y externo. En la cara nuclear convergen para formar una canastilla o cesta. A lo largo de estas fibrillas se ubican nucleoporinas que intervienen en el transporte de sustancias a través del poro.
· Un poro central o abertura.
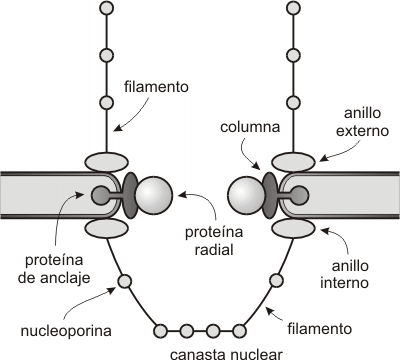
Fig. 10.4 - Esquema del complejo de poro nuclear
La luz de los CPN suele presentarse obturada por las proteínas que circulan a través del poro, como por las carioportinas (Kap) que actúan como eficientes transportadores en el tráfico núcleo/citoplasma.
Los CPN presentan uno o varios canales acuosos a través de los cuales las pequeñas moléculas solubles en agua difunden (transporte no regulado). Las moléculas de mayor peso molecular son transportadas en forma activa, por lo que requieren energía y moléculas transportadoras.
Se importan dentro del núcleo:
· Las proteínas sintetizadas en el citoplasma necesarias para ensamblar los ribosomas.· Los factores de transcripción requeridos en la activación o inactivación de los genes.
· Los factores de empalme necesarios en el proceso de maduración de los ribosomas.
Las moléculas y macromoléculas ensambladas y exportadas desde el núcleo al citoplasma incluyen:
· Las subunidades ribosomales
· ARNm
· ARN de transferencia
· Factores de transcripción que son devueltos al citoplasma para ser reutilizados.
Los mecanismos implicados en el transporte a través del poro son diferentes al transporte de proteínas en las membranas de otros organelos . Por ejemplo, las proteínas nucleares son transportadas a través del poro manteniendo su conformación plegada, por el contrario las proteínas que no se localizarán en el núcleo se despliegan durante el transporte.
Los complejos de poro nuclear hacen de la envoltura nuclear una barrera selectiva entre el núcleo y el citoplasma. Estos complejos constituyen la principal vía de comunicación entre el compartimiento nuclear y citoplasmático de la célula ante el pesado tráfico molecular. Aun cuando las proteínas pequeñas y otras moléculas viajan a través de los canales periféricos, las proteínas de gran tamaño deben poseer una etiqueta para ingresar por el canal central. Estas proteínas sintetizadas en el citoplasma contienen la señal de localización nuclear (nuclear signal localization, NSL).
Tampoco los ARN pueden salir de núcleo por sí mismos. Ellos salen a través del complejo de poro con una proteína especial que posee una señal nuclear de exportación (nuclear export signal, NES). Ambas NSL y NES consisten en una secuencia corta de aminoácidos, muchos de los cuales tienden a estar ubicados centralmente dentro de las proteína.
Observemos en la Fig. 10.4, como las proyecciones filamentosas desde la cara citosólica del complejo pueden enlazar proteínas, conduciéndolas dentro del poro central. Los filamentos citosólicos, el poro central y la canasta nuclear se proyectan dentro del compartimiento nuclear. Se cree que la canasta puede ser un área importante de paso para la preparación de las ribonucleoproteínas (RNP) antes de ser exportadas.
Las moléculas de mayor tamaño requieren de una proteína “transbordadora” o carioportina. La superfamilia de carioportinas esta integrada por: importinas . exportinas y transportinas .
ImportaciÓn de proteÍnas (Fig. 10.5)
Las importinas son heterodímeros, formados por dos subunidades, la subunidad-a se une a la NSL de la proteína nuclear permitiendo la unión con la subunidadad-b. Esta unión origina una “importina funcional” que lleva unida a la proteína nuclear a ser transportada.
El complejo importina funcional se pone en contacto con los filamentos citosólicos, donde guiado por las nucleoporinas (Nup), llega al poro central. La translocación de complejo importina/carga es regulado por la pequeña RanGTPasa [1] , que se une a la subunidad b de la importina. Esta b-importina es la encargada de interactuar con el poro provocando su dilatación y posibilitando el ingreso de la proteína nuclear. La translocación de proteínas es un proceso activo. Cuando el complejo penetra al interior del núcleo, las subunidades de importina se separan y la carga es liberada. La disociación de las subunidades causa entonces un nuevo cambio en su forma, dejando al descubierto la NES de cada subunidad. Otras proteínas en el poro central reconocen la NES y retornan las subunidades al citoplasma.
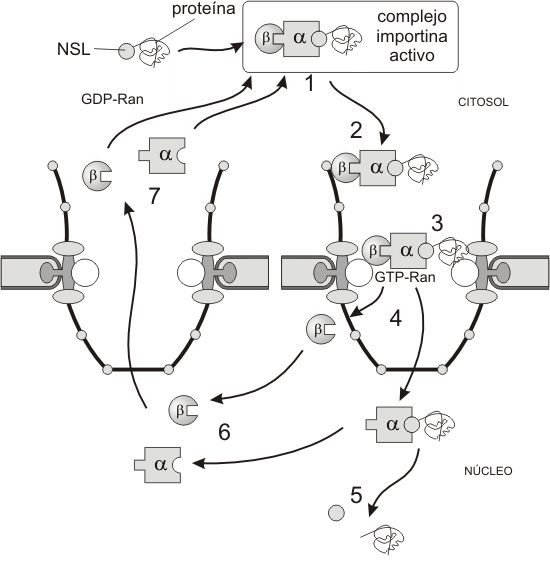
Fig. 10.5 - Modelo de mecanismo de importación a través del CPN
ExportaciÓn de ARN
Los ARN maduros se asocian a proteínas llamadas transportinas, las cuales actúan como transbordadores permitiendo el pasaje de ARN al citoplasma. En la fig. 10.6 se esquematiza como el ARNm es llevado fuera del núcleo. Los ARNm maduros que presentan la poli A se asocian con varias proteínas, formando una partícula de ribonucleoproteína (RNP). Estas partículas se mueven linealmente a través de la canasta nuclear. Al igual que las importinas, las RNP son recicladas hacia el núcleo. En el citoplasma, las CRBP ( del inglés, cytoplasmic RNA-binding proteins) reemplazan a las RNP para guiar a los ARNs a sus destinos citosólicos correctos.
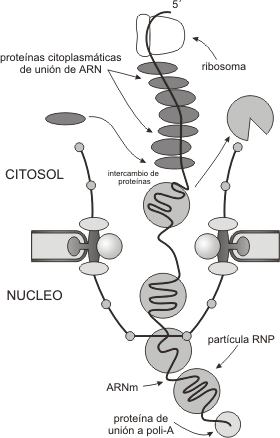
Fig. 10.6 - Modelo de mecanismo de exportación de ARNm a través del CPN
Cromosomas y Cromatina
El núcleo contiene los cromosomas de la célula. Cada cromosoma consiste en una molécula única de ADN con una cantidad equivalente de proteínas. Colectivamente, el ADN con sus proteínas asociadas se denomina cromatina. La mayor parte de las proteínas de la cromatina consisten en copias múltiples de cinco clases de histonas.
Estas proteínas básicas son ricas en residuos de arginina y lisina cargados positivamente. Por esta razón se unen estrechamente con los grupos fosfatos (cargados negativamente) del ADN.
La cromatina también contiene pequeñas cantidades de una amplia variedad de proteínas no histónicas y RNP. La mayoría de ellas son factores de transcripción (por ej., el receptor esteroide), siendo su asociación con el ADN pasajera. Estos factores regulan que parte del ADN será transcripta en ARN.
Niveles de organizaciÓn de la cromatina
La observación a través del microscopio óptico de un núcleo interfásico, nos permite distinguir dos tipos de cromatina. La eucromatina o cromatina laxa, de localización central, y la heterocromatina o cromatina densa, en la periferia del núcleo (Fig.10.7). La heterocromatina representa aproximadamente el 10% del total de cromatina y es considerada transcripcionalmente inactiva (Fig. 10.8).
La eucromatina se encontraría al menos en dos estados, la eucromatina accesible, que representa alrededor del 10%, donde se encuentran los genes que se están transcribiendo y la eucromatina poco accesible, más condensada (pero menos que la heterocromatina), donde están los genes que la célula no está transcribiendo.Si el núcleo celular se incuba con nucleasas, enzimas que digieren el ADN, las secuencias que primero se digieren son las que portan los genes expresados por la célula, lo que corrobora el menor grado de condensación de la eucromatina.
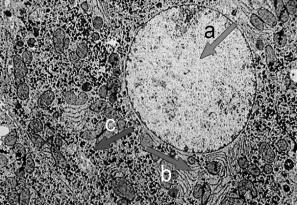
Fig. 10.7 - Microfotografía electrónica del núcleo de un hepatocito. a. eucromatina; b. RER

Fig. 10.8 - Microfotografía
electrónica del núcleo de un linfocito. a. eucromatina; b. heterocromatina,
c. mitocondria
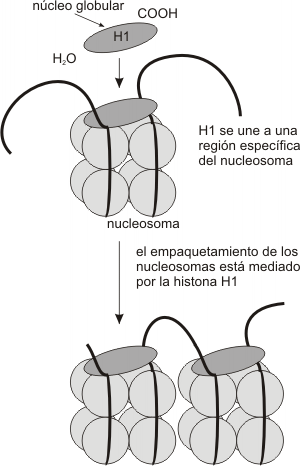
Fig. 10.9 - H1 y formación de la fibra de 10 nm
Cuando el cromosoma en interfase se esparce artificialmente sobre agua, tiene la apariencia de un collar de perlas. Las perlas son los nucleosomas, las unidades de enrollamiento de la cromatina. (Fig. 10.10b)
Los nucleosomas están formados por un centro o "core" de histonas. Dicho centro posee dos copias de cada una de las siguientes histonas: H2A; H2B; H3 y H4
Alrededor del centro de histonas, 146 pares de bases del ADN se enrollan en dos vueltas. La unión de las histonas al ADN no depende de una secuencia particular de nucleótidos, sino de la secuencia de aminoácidos de la histona. Las histonas son unas de las moléculas más conservadas durante el transcurso de la evolución. La histona H4 en el ternero difiere de la H4 de la planta de poroto en sólo 2 residuos de aminoácidos de una cadena de 102. Alrededor de 60 pares de bases de ADN unen un nucleosoma con el próximo. Cada región de unión es el ADN espaciador. La quinta histona, la H1, conecta a los nucleosomas y actúa como una banda de goma, manteniéndolos juntos dentro de una misma cuerda enrollada. Esta estructura se conoce como fibra de 10nm, siendo el primer grado del empaquetamiento de la cromatina. (Fig. 10.9)Los nucleosomas se organizan, a su vez, en fibras de 30nm (solenoide), girando a manera de resorte alrededor de un eje virtual. Esta estructura es mantenida por la interacción de las H1 de nucleosomas cercanos. (Fig. 10.10c)
En el siguiente nivel de empaquetamiento, las fibras de 30 nm se organizan en una serie de bucles o asas superenrolladas (Fig. 10.10d; Fig. 10.11). Estos bucles se estabilizan gracias a la interacción con las proteínas de la matriz nuclear o andamiaje nuclear (“scaffold”).
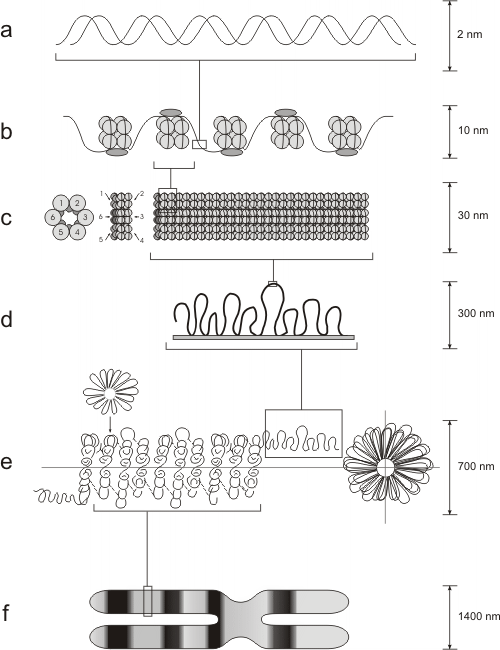
Fig. 10.10 - Modelo de empaquetamiento de la cromatina
Cada bucle de cromatina representa un dominio funcional o unidad de replicación (Fig. 10.10e). Estos dominios contienen alrededor de 100.000 pares de bases, extensión de ADN suficiente para acomodar varios genes de tamaño promedio. Algunos genes, sin embargo, pueden abarcar varios dominios adyacentes de un cromosoma. Cada cromosoma puede tener cien o más dominios.Durante la profase, los cromosomas aparecen en forma más condensada, alcanzando la cromatina su mayor nivel de condensación en metafase(Fig. 10.10f). La organización de los cromosomas envuelve la fosforilación de la H1 y otras proteínas, lo cual causa el plegamiento y empaquetamiento aún más compacto de la cromatina. El andamiaje o matriz nuclear se convierte en el centro de la estructura del cromosoma, y como la compactación continúa, éste se pliega modo de acordeón.
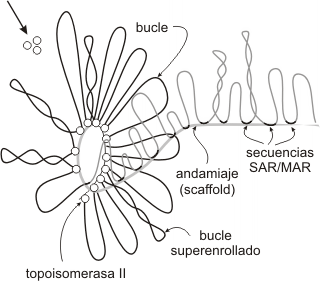
Fig. 10.11- Empaquetamiento de la fibra de 30 nm
El grado de condensación de los dominios de cromatina se mantiene principalmente debido a la asociación con la matriz nuclear y a proteínas asociadas como la topoisomerasa II o girasa, encargada de controlar el grado de superenrollamiento del ADN (Fig. 10.11). La unión entre la cromatina y la matriz se da a nivel de zonas altamente conservadas, denominadas secuencias SAR o MAR (scaffold associated regions/ matrix attachment regions). Las SAR son regiones de varios cientos de pares de bases ricas en residuos de adenina y timina, abundantes en la heterocromatina.
Con coloraciones especiales los cromosomas, revelan diferencias estructurales de importancia funcional. Las bandas oscuras consisten en cromatina altamente condensada, mientras que las bandas claras se corresponden con cromatina más laxa.
El examen de la cromatina en bandas claras y oscuras revela que ambos tipos de cromatina están acomodados en bucles de distintos tamaños y que a su vez se proyectan desde el andamiaje plegado. El andamiaje está muy plegado en la heterocromatina, y es más lineal en las bandas de eucromatina, formando bucles más amplios. Las histonas de la eucromatina están fuertemente acetiladas. Estos cambios afectarían el grado de empaquetamiento de la eucromatina, haciéndola más accesible para la transcripción de sus genes.
Las características de la hetero y eucromatina son:
|
Tabla 10.1 - Características de la Cromatina |
||||
|
Tipo de cromatina |
Estado físico |
Cambio químico |
Tipo de genes |
Replicación |
|
Eucromatina |
Laxa |
Acetilada |
Activos |
Fase S temprana |
|
Heterocromatina |
condensada |
Metilada |
Silentes |
Fase S tardía |
Los cromosoma en metafase también poseen un revestimiento de RNP. Dicho revestimiento deriva de los componentes del nucléolo. Estos cromosomas se constituyen en el vehículo para dividir el material nucleolar entre las futuras células hijas. El empaquetamiento de la cromatina permite confinar al ADN dentro del núcleo La molécula de ADN de un cromosoma humano contiene 50 x 106 pares de nucleótidos en el cromosoma más pequeño (1.7 cm con la molécula extendida) a 250 x 106 pares de nucleótidos en el más largo ( 8.5 cm). Midiendo extremo con extremo el total de cromosomas de una célula humana diploide, el ADN se extiende más de 2 metros. El empaquetamiento del ADN en forma de cromatina, no solamente le permite entrar dentro de los límites del núcleo, sino también lo protege del ataque de las nucleasas.
El cromosoma eucariota (Fig. 10.12)
Cada cromosoma eucariota consiste en una molécula simple de ADN de alrededor de 150 millones de pares de nucleótidos.La molécula de ADN en el cromosoma eucariota es lineal, por lo tanto posee dos extremos (en contraste con el cromosoma bacteriano que es circular).
La molécula de ADN de un cromosoma típico eucariota contiene:
· Un conjunto lineal de genes que codifican para ARN y proteínas interrumpido por
· Muchas secuencias de ADN no codificante.
El ADN no codificante incluye:
· Secuencias de aproximadamente 170 nucleótidos de ADN satélite, repetidas miles de veces, que corresponden al centrómero.
· Secuencias repetitivas en los extremos del cromosoma llamadas telómeros.
· Múltiples secuencias señalizadoras altamente conservadas, denominadas origen de replicación (ORI) , necesarias para que se realice la duplicación del ADN en un tiempo breve.
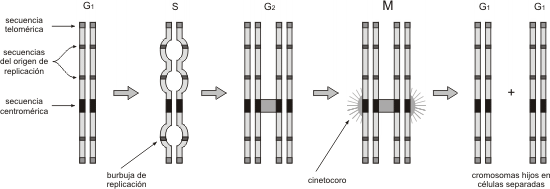
Fig. 10.12- Secuencias de un cromosoma eucariota estable en las diferentes etapas del ciclo celular
El centrómero es una constricción primaria localizada centralmente o hacia los extremos de cada cromosoma.
El ADN centromérico como ya mencionamos es altamente repetitivo y se encuentra siempre condensado siendo parte de la heterocromatina.
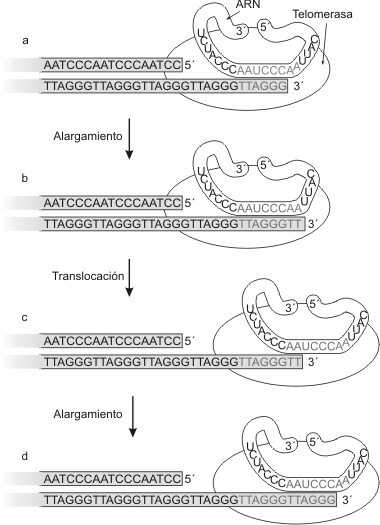
Fig. 10.13- Síntesis de telómeros por la telomerasa. a) La telomerasa se une al telomero; b) La telomerasa alarga los extremos de la cadena 3`; c) La telomerasa avanza; d) La ADN polimerasa sintetiza la cadena retrasada.
Los telómeros son cruciales en la vida de la célula. Ellos son necesarios para la duplicación completa del cromosoma, los protegen de las nucleasas, evitan que los extremos del cromosoma se fusionen entre sí y facilitan la interacción del cromosoma con la envoltura nuclear.
Los telómeros de las células humanas contienen la secuencia 5'TTAGGG3, que se repite aproximadamente 2000 veces.5 '... ..TTAGGG TTAGGG TTAGGG TTAGGG TTAGGG TTAGGG..... 3 '
3 '... ..AATCCC AATCCC AATCCC AATCCC A..... 5 '
La cadena rica en guanosina corre en dirección 5' a 3', extendiéndose 12 a 15 nucleótidos más allá de la cadena rica en citosina, formando un apéndice en una de las cadena en cada extremo del cromosoma. Este desnivel se mantiene de generación en generación por medio de una enzima especial, la telomerasa, que agrega nuevas unidades al extremo 3' de la cadena rica en guanosina (Fig. 11.13).La telomerasa es una ribonucleoproteína, la cual provee un molde de AAUCCC que guía la inserción de la secuencia TTAGGG. Entonces la telomerasa es una retrotranscriptasa, sintetiza ADN a partir de un molde de ARN.
Las células con telomerasa activa pueden compensar el acortamiento de los telómeros durante la duplicación del ADN.
La telomerasa activa se encuentra solamente en:
· Las células de la línea germinal, incluyendo células troncales embrionarias
· Eucariotas unicelulares
· Células cancerosas
Los cromosomas se diferencian por la ubicación del centrómero

Fig. 10.14- Microfotografía electrónica de un cromosoma metafásico
Antes de que una célula se divida, cada cromosoma se duplica (durante la fase S del ciclo celular).
Al inicio de la división celular, los cromosomas duplicados se condensan en estructuras que pueden teñirse con facilidad (por ello denominadas cromosomas), pudiéndose observar bajo el MO.
La condensación es tal que el cromosoma es aproximadamente 10.000 veces más corto que la molécula de ADN que contiene (Fig. 10.14). A primera vista, los cromosomas duplicados se mantienen juntos por el centrómero. Mientras están juntos, es común llamar a cada parte del cromosoma duplicado, cromátida hermana.
Esto no debe confundirnos, cada una de las "cromátidas hermanas" es un cromosoma completo. El cinetocoro es una estructura proteica discoidal que forma parte del centrómero y ayuda a separar las cromátidas hermanas. Es el sitio de unión con los microtúbulos del huso, que contienen los motores de dineína que tiran a los cromosomas en la anafase. Además proveen una plataforma para ensamblar y movilizar las proteínas que construyen el huso.
La posición del centrómero, determina el largo de los brazos del cromosoma; en base a esto se puede clasificar a los cromosomas en: (Fig. 10.15)
· Metacéntricos: el centrómero en posición central determina brazos de igual longitud
· Submetacéntricos: un par de brazos es más corto que el otro, pues el centrómero se encuentra alejado del centro.
· Acrocéntricos: el centrómero se halla próximo a uno de los extremos, por lo tanto uno de los brazos es casi inexistente.
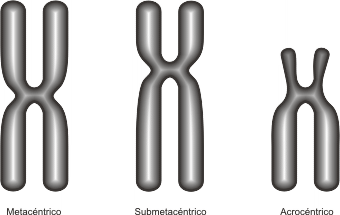
Fig. 10.15- Tipos de cromosomas
Los cromosomas acrocéntricos poseen una masa de cromatina llamada satélite, en el extremo del brazo corto. El satélite se halla aislado del resto del cromosoma por la constricción secundaria. La zona aledaña al satélite de los cromosomas acrocéntricos contribuye a formar el nucléolo (Fig. 10.16)
El más corto de los dos brazos del cromosoma se llama p; el más largo es el brazo q.
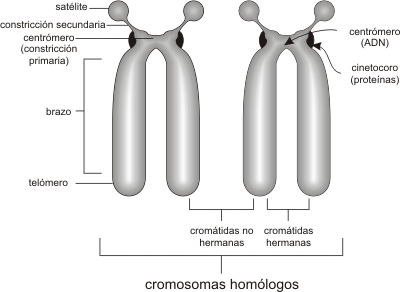
Fig. 10.16- Partes de un cromosoma mitótico.
Todas las especies tienen un número característico de pares de cromosomas homólogos llamado número diploide (2n). El número diploide del hombre es 46.
El cariotipo es una representación gráfica o fotográfica de los cromosomas presentes en el núcleo de una sola célula somática de un individuo. Cada miembro del par de cromosomas homólogos proviene de cada uno de los padres del individuo cuyas células examinamos.
El cariotipo de la mujer contiene 23 pares de cromosomas homólogos, 22 pares son autosomas y el par restante, cromosomas sexuales, ambos " X".
El cariotipo del hombre contiene los mismos 22 pares de autosomas y 1 par de cromosomas sexuales, un cromosoma sexual "X" y un cromosoma sexual "Y" (un gen en el cromosoma Y designado SRY es el que pone en marcha el desarrollo de un varón, por lo tanto determina el sexo).
El análisis del cariotipo involucra la comparación de cromosomas por su longitud, la ubicación de los centrómeros y la ubicación y los tamaños de las bandas G.
Durante la mitosis, los 23 pares de cromosomas humanos se condensan y son visibles con un microscopio óptico.
Preparación de un Cariotipo: La preparación de un cariotipo normalmente involucra bloquear las células (glóbulos blancos) durante la mitosis con colchicina y marcar los cromosomas condensados con tinción Giemsa. La tinción marca las regiones de los cromosomas que son ricos en pares de nucleótidos entre A -T produciendo una banda oscura, la banda G. Luego de la tinción, los cromosomas se fotografían, se recortan y se ordenan de acuerdo a su longitud. Los de igual tamaño se aparean según la ubicación de su centrómero.
Una error común es suponer que cada banda representa un sólo gen. En realidad las bandas más pequeñas contienen más de un millón de pares de nucleótidos y potencialmente cientos de genes. Por ejemplo, el tamaño de una banda pequeña es igual a toda la información genética de una bacteria.
El análisis del cariotipo es una de muchas técnicas que nos permiten investigar las miles de enfermedades genéticas que se pueden encontrar en los seres humanos.
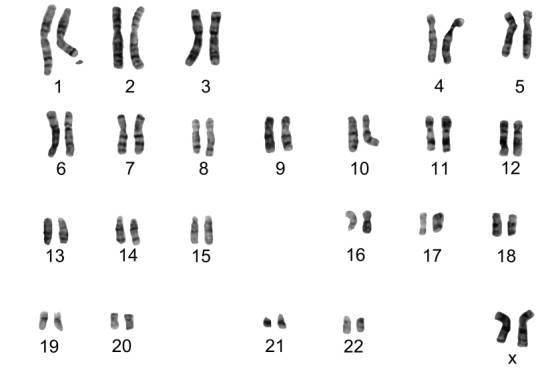
Fig. 10.17 - Cariotipo femenino normal: Los autosomas se ordenan en grupos por tamaño y posición del centrómero.
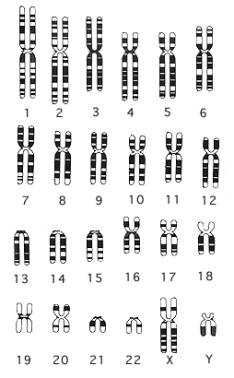 |
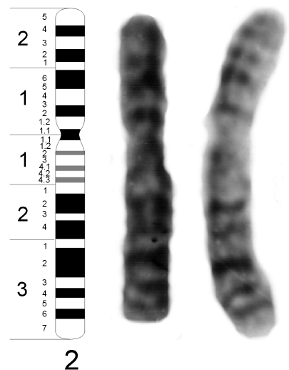 |
Fig. 10.18 - Mapa estándar (Idiograma) de patrones de bandeo del cromosoma 2. Los números corresponden a las regiones y bandas. A la derecha , idiograma del cariotipo humano masculino.
El nucléolo
En el nucléolo tiene lugar la formación de subunidades ribosómicas, la síntesis y procesamiento de ARNr y actualmente se considera que desempeña un importante papel en la regulación del ciclo celular. [2]
El nucléolo es un aglomerado de fibras de cromatina de distintos cromosomas. En el hombre, los pares 13,14, 15, 21 y 22, aportan sectores de cromatina que forman el nucléolo. Todos estos cromosomas son acrocéntricos y presentan constricciones secundarias denominadas organizadores nucleolares (NOR), donde están los genes que codifican ARNr (Fig. 10.19).
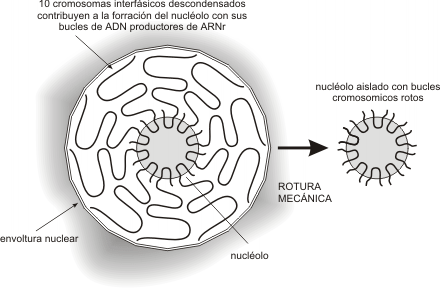
Fig. 10.19 - Esquema de nucléolo indicando los bucles de los 10 cromosomas con los genes para el ARNr
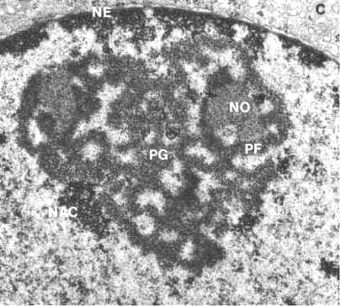
Fig. 10.20 - Microfotografía electrónica del nucléolo. NE, Envoltura nuclear; NO. Organizador nucleolar; PG, Zona granular; PF, Zona fibrilar.
El nucléolo aparece como una estructura simple carente de componente membranoso, en la que diferenciamos dos regiones:
· Una zona fibrilar central, formada por ADNribosómico y ARNr naciente
· Un zona granular periférica donde los gránulos están formados por las subunidades ribosómicas en proceso de ensamblado (Fig. 10.20).
Los nucléolos, al igual que la envoltura nuclear desaparecen en la mitosis y se reorganizan alrededor de los segmentos de ADNr, que como su nombre lo indica, codifica ARNr. Siendo el ARNr el más abundante dentro de los tipos de ARN, existen múltiples copias del gen que lo codifica. El genoma humano presenta alrededor de 200 copias del gen para ARNr. Estos genes que promedian los 10.000 nucleótidos se localizan en tándem. Cada gen está separado por ADN espaciador y presenta asociado una molécula de ARN polimerasa I. De cada enzima parten perpendicularmente los ARNr nacientes, tomando la apariencia característica de un árbol de navidad. Cada gen produce un transcripto llamado ARNr 45S que será luego procesado (Fig. 10.21)
El tamaño del nucleólo varía entre células y en la misma célula según su actividad, pues si bien la velocidad de transcripción puede acelerarse, el ensamblado de las subunidades ribosomales requiere de un tiempo más o menos constante; es por ello que en los nucléolos grandes observamos mayor proporción de componente granular.
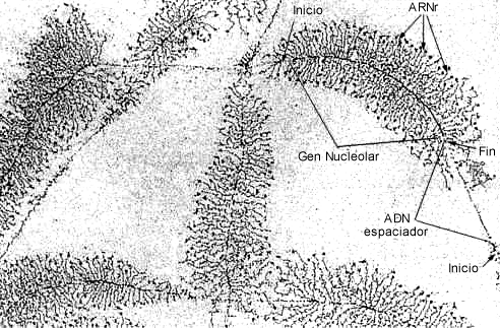
Fig. 10.21 - Microfotografía electrónica de los genes nucleolares (ADNr) durante la transcripción. Observe como la longitud de los transcriptos primarios aumenta a medida que nos alejamos del punto de inicio.
AutoevaluaciÓn
1) La eucromatina se caracteriza por:
a- ser transcripcionamente inactiva
b- teñirse debilmente
c- presentarse altamente condensada
d- formar parte de genes que no se expresan
2) En el nucléolo se sintetizan:
a- Precursores de ARNr
b- ARNt
c- Pre ARNm
d- ARNm
3) Los organizadores nucleolares aportan información para la síntesis de:
a- ARNm
b- ARNr
c- ARNt
d- Ribosomas
4) La composición química y función de las histonas son respectivamente:
a- proteínas básicas y forman parte en la estructura de la cromatina
b- proteínas básicas cuya única función es regular la expresión de los genes
c- son proteínas ácidas e intervienen en la estructura de la cromatina
d- son proteínas ácidas e intervienen en la síntesis de ADN polimerasa.
5) El andamiaje o matriz nuclear:
a- está formado exclusivamente por láminas
b- se asocia a la cromatina a nivel de las SAR/MAR del ADN
c- está más replegado en la eucromatina
d- se desorganiza durante la división celular
6) Presentan señal de localización nuclear:
a- las hormonas esteroides
b- todas las riboproteínas
c- los factores de transcripción
d- todas las anteriores son correctas
7) Los dominios funcionales de la cromatina:
a- aparecen por empaquetamiento de la fibra de 10 nm
b- dependen exclusivamente de la interacción con la H1
c- representan individualmente un gen
d- funcionan como unidades de replicación del cromosoma.
8) El pasaje de moléculas a través de los CPN:
a- no es regulado y requiere de etiquetas o señales en las moléculas transportadas
b- no es regulado para moléculas pequeñas y solubles que circulan por los canales periféricos
c- siempre es regulado y no depende del tamaño de la molécula a transportar
d- requiere que todas las moléculas a ser transportadas tengan una NSL
9) ¿Cuál de las siguientes afirmaciones es FALSA con respecto a la telomerasa?
a- es la enzima encargada de formar los telómeros
b- es una ARN polimerasa
c- se inactiva poco después del nacimiento
d- las células cancerosas conservan la telomerasa siempre activa
10) Un investigador inyecto un cultivo celular con suero anti-laminina, y observó:
a- que la células no pudieron dividirse
b- que las células luego de la mitosis no pudieron organizar su envoltura nuclear
c- que los complejos laminina/antilamina se acumularon dentro del núcleo
d- que la envoltura nuclear formada resulto frágil e inestable
Bibliografía
- Alberts, B et al. (2010). Biología Molecular de la Célula. 5ta. Edición. Ediciones Omega S.A. Barcelona.
- Becker W, Kleinsmith L y Hardin J (2007). El mundo de la célula. 6ta. Edición. Pearson Educación. Madrid.
- Cooper G, Hausman R (2010). La Célula. 5ta edición. Marbán Libros. Madrid.
- Curtis H, Schnek A, Massarini A. (2015). Invitación a la Biología en contexto social. 7ma. Edición. Editorial Médica
Panamericana. Bs. As.
- De Robertis(h), Hib J (2012). Biología Celular y Molecular de De Robertis. 16ma. Edición. Ediciones Promed. Bs.As.
- Karp, G. (2014). Biología Celular y Molecular. 6ta. Edición.Ed. Mc Graw Hill Interamericana. México
- Lodish H et al (2016). Biología Celular y Molecular. 7ma. Edición. Editorial Médica Panamericana. Bs. As.
- Pierce B.(2016). Genética. 5ta. Edición. Editorial Médica Panamericana. México.
[1] En el nucléolo se secuestran algunas de las proteínas reguladoras del ciclo celular como la Cdc14, durante G1, S, G2 y metafase.
[2] Las Ran GTPasas nucleares, no sólo intervienen en el transporte nuclear, también son requeridas durante el procesamiento de ARN, el ensamblado del aparato mitótico, la descondensación de los cromosomas y el crecimiento nuclear luego de la mitosis.