COMUNICACIÓN INTERCELULAR Y TRANSMISIÓN DE SEÑALES
Silvia Márquez – Lionel Valenzuela Pérez – Sergio D. Ifrán – Maria Elena Pinto – Gladys Gálvez
Bases Molecular de la comunicación Intercelular
Los organismos unicelulares pueden realizar todas las funciones necesarias para mantener la vida. Por ejemplo, una ameba, organismo unicelular, asimila los nutrientes del medio, se mueve, lleva a cabo las reacciones metabólicas de síntesis y degradación y se reproduce. En los organismos pluricelulares, la situación es mucho más compleja, ya que las diversas funciones celulares se distribuyen entre distintas poblaciones de células , tejidos y órganos. De este modo en un organismo pluricelular, cada célula depende de otras y las influye. Por lo tanto la mayoría de las actividades celulares, solo se desarrollan, si las células involucradas son alcanzadas por estímulos provenientes de otras. Para coordinar todas estas diversas funciones deben existir mecanismos de comunicación intercelular.
Cuando una célula recibe un estímulo puede responder con alguno de los siguientes cambios, dependiendo de las características del estímulo y el tipo de célula receptora del mismo: por ejemplo, se puede diferenciar, reproducir, incorporar o degradar nutrientes, sintetizar, secretar o almacenar distintas sustancias, contraerse, propagar señales o morir.
Inducción
En la mayoría de los organismos superiores existen dos métodos fundamentales de comunicación intercelular: un sistema fundado en las neuronas o células nerviosas y otro basado en las hormonas. En ambos sistemas las células se comunican entre si a través de mensajeros químicos.
Las neuronas envían mensajes a sus células efectoras (células blanco), que pueden ser células musculares, células glandulares u otras neuronas. Para enviar su mensaje, la neurona libera una sustancia química, un neurotransmisor. El neurotransmisor es liberado en sitios específicos llamados sinapsis [1] . Las moléculas de neurotransmisor se unen a receptores, situados en la superficie de la célula blanco, y provocan de esta forma cambios físicos y químicos en la membrana celular y en el interior celular.
Por lo tanto diremos que en general, la acción de estimular a las células desde el exterior se llama inducción y se realiza a través de sustancias producidas por células inductoras. La célula que es sensible al inductor se denomina célula inducida, blanco o diana y presenta para el mismo receptores específicos (fig. 7.1), que pueden ubicarse en la membrana plasmática, el citoplasma o en el núcleo. Estos receptores son proteínas o complejos proteicos.
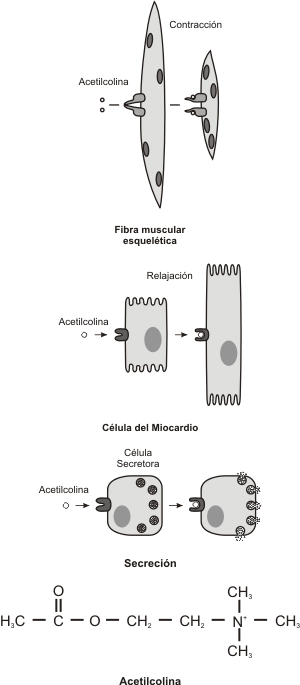
Fig. 7.1- Efecto de un mismo inductor sobre diferentes células blanco. Un inductor puede tener varios receptores, causando distintas respuestas celulares
Cuando el receptor se encuentra en el citoplasma o en el núcleo, el inductor debe ser pequeño e hidrófobo, de modo que pueda atravesar la membrana plasmática sin dificultad, mientras que los receptores de membrana pueden recibir inductores de cualquier tipo.
La acción de las hormonas, puede darse básicamente de acuerdo a uno de estos cinco tipos de inducción:
1. Endocrina: una glándula libera hormonas (inductor) que pueden actuar sobre células u órganos situados en cualquier lugar del cuerpo (células blanco). Por lo tanto podemos decir que células inductoras e inducidas se encuentran distantes. Las glándulas endocrinas liberan hormonas al torrente sanguíneo: las células o tejidos blanco poseen receptores que reconocen exclusivamente los diferentes tipos de moléculas hormonales. Así un receptor reconoce exclusivamente una hormona. Una célula puede tener distintos tipos de receptores, y así reconocer diferentes hormonas. Ej. Insulina, glucagón, hormonas adenohipofisiarias, etc.
2. Paracrina: Una célula o un grupo de ellas liberan una hormona que actúa sobre las células adyacente que presenten el receptor adecuado. De esta forma la célula inductora e inducida se encuentran próximas. Ej. Prostaglandinas
3. Autocrina: Una célula libera una hormona que actúa sobre la misma célula. Ej. prostaglandinas
4. Neuroendocrina: Una neurona libera su neurosecreción al torrente sanguíneo. Ej. Oxitocina, ADH, hormonas liberadoras e inhibidoras hipotalámicas
5. Por contacto directo: La hormona o molécula inductora es retenida en la membrana plasmática de la célula inductora, por lo tanto no se secreta. Las células deben ponerse en contacto, para que la sustancia inductora tome contacto con el receptor localizado en la membrana plasmática de la célula inducida. Ejemplo de este tipo de comunicación tienen lugar en algunas respuestas inmunológicas.
6. Yuxtacrina ( a través de uniones comunicantes, nexus o gap: Las células conectadas a través del establecimiento de este tipo de uniones firmes, puede responder de forma coordinada ante un inductor que se une a alguna de las células que están comunicadas. A través de estas uniones pasan pequeñas moléculas como los segundos mensajeros.
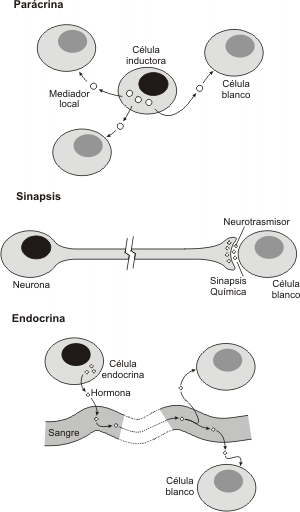
Fig. 7.2 - Algunas formas de inducción por moléculas secretadas
Como vemos existen importantes diferencias entre la comunicación hormonal y la nerviosa. Las neuronas tienden a actuar sobre una célula en particular o sobre un grupo de ellas. Generalmente los axones recorren distancias cortas , aunque existen excepciones a esta regla. La comunicación entre neuronas puede desarrollarse en cuestión de milisegundos. Por el contrario, una hormona liberada al torrente sanguíneo por una glándula, puede alcanzar células y tejidos en cualquier parte del cuerpo, siempre que estas tengan el receptor adecuado, además la comunicación hormonal puede prolongarse por espacio de minutos o varias horas.
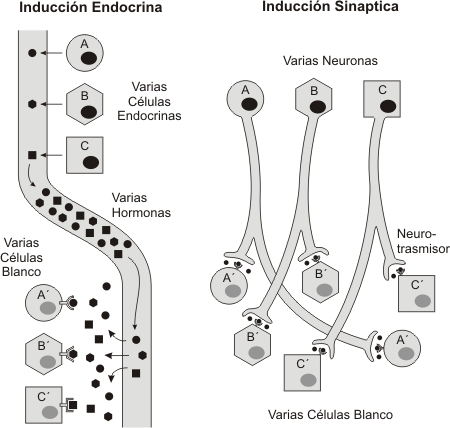
Fig. 7.4 - Inducción endócrina versus inducción sináptica. Observe como la hormona vehiculizada por la sangre alcanza a todas las células del cuerpo, uniendose sólo a las que presentan receptores específicos. En la sinapsis, el neurotransmisor transportado a las terminales nerviosas por flujo axónico, es liberado en el espacio sináptico, alcanzando sólo a las células efectoras próximas a la terminal nerviosa.
Características del complejo inductor- receptor
Cuando una hormona pasa a la circulación sanguínea, puede alcanzar todos los tejidos del cuerpo, sin embargo, por lo general su acción sólo se evidencia en un limitado número de células. Como señaláramos, el receptor es por lo general un complejo proteico específico al que cada inductor se une selectivamente, de este modo la sustancia inductora y su receptor forman un complejo que presenta las siguientes características:
Encaje inducido: La unión inductor- receptor supone una adaptación estructural entre ambas moléculas, similar al complejo enzima-sustrato.
Saturabilidad: ya que el número de receptores en una célula es limitado, un eventual aumento en las concentraciones del inductor, pondría en evidencia la saturabilidad del sistema.
Reversibilidad: El complejo inductor-receptor se disocia después de su formación.
La interacción inductor-receptor es la primera de una serie de reacciones consecutivas
que se propagan por el interior de la célula, mientras que el último eslabón de esta serie puede considerarse cómo la respuesta.
Como ya lo adelantáramos y de acuerdo a la ubicación de los receptores específico, los inductores se pueden clasificar en dos grupos: a) los que se unen a receptores de membrana y b) los que ingresan a la célula y se unen a receptores citosólico.
A su vez las moléculas que actúan como hormonas pueden clasificarse de acuerdo a su estructura química en cuatro categorías:
1. Esteroides: Las hormonas esteroides son derivados del colesterol. Ejemplos de las hormonas esteroides son los glucocorticoides, los mineralocorticoides, los esteroides sexuales, la vitamina D y el ácido retinoico.
2. Derivados de aminoácidos: hormonas derivadas del aminoácido tirosina. Conocidas como aminohormonas. Existen dos tipos de aminohormonas las que interactúan con receptores de membrana (adrenalina y noradrenalina, producidas por la glándula suprarrenal) y las que se unen a receptores citosólicos (por ejemplo, la hormona tiroidea producida por la glándula tiroides).
3. Péptidos o proteínas: Son cadenas de aminoácidos. Ejemplos de hormonas peptídicas son la oxitocina y la hormona antidiurética. Ejemplos de hormonas proteicas son la Insulina y la hormona del crecimiento. Estas proteínas y otros factores de crecimiento son mitógenos potentes. (es decir activan la mitosis).
4. Derivados de ácidos grasos: Las prostaglandinas y las hormonas juveniles de los insectos son hormonas derivadas de ácidos grasos.
Debemos recordar que estas moléculas son mensajeros químicos, cuya función es coordinar las respuestas de las distintas poblaciones celulares en un organismo pluricelular. Sin embargo, estos mensajeros químicos no actúan de la misma forma. Por ejemplo las hormonas peptídicas y proteicas debido a su tamaño y polaridad, no pueden atravesar la membrana plasmática y deben unirse a receptores dispersos en la superficie externa de la célula. Estos son los llamados receptores de membrana, que en general son glicoproteicos. Los receptores de membrana detectan la llegada de una hormona y activan una ruta de transmisión de señales intracelular, que en ultima instancia regula los procesos celulares. Por lo tanto en este caso podemos decir, que la membrana plasmática celular constituye una barrera que se opone al flujo de información. En la membrana plasmática se alojan mecanismos que transducen las señales externas, en otras internas, responsables últimos de la regulación de las funciones celulares. En general vamos a denominar a las señales externas (hormonas), como primeros mensajeros, y a las señales internas como segundos mensajeros. El proceso de generar los segundos mensajeros, depende de una serie de proteínas de la membrana celular. Los segundos mensajeros son en general moléculas de pequeño tamaño, cuya rápida difusión permite que la señal se propague rápidamente por todo el interior celular.
El otro tipo de señales extracelulares (inductores) son las hormonas esteroideas y las hormonas tiroideas, que por su naturaleza hidrofóbica (liposoluble), pueden difundir a través de la membrana plasmática, e interactuar directamente con receptores que se encuentran en el interior de la célula, por ejemplo en el citosol . Una vez que el inductor, interactua con el receptor citosólico, formando un complejo Hormona-Receptor, este complejo ingresa al núcleo donde activan genes específicos.
Base molecular de la comunicaciÓn intracelular
Inducciones celulares mediadas por receptores de membrana asociados a proteínas G
Podemos decir que las rutas de transmisión de información intracelular comparten una secuencia de procesos. Los mensajeros externos (primer mensajero), se unen a las moléculas receptoras que activan a las proteínas transductoras asociadas al receptor. Estas proteínas una vez activadas, transportan señales a través de la membrana a las enzimas amplificadoras, que generan las señales internas transportadas por los segundos mensajeros.
En este caso de inducción, el receptor de membrana, transmite la información a través de la membrana plasmática, hacia el interior de la célula, por medio de una proteína transductora, la proteína G. Las proteínas G poseen tres subunidades, alfa, beta y gamma. La subunidad alfa puede unir GTP y también puede degradarlo (actividad GTPasa). El dímero beta-gamma mantiene a la proteína G unida a la membrana. Estas proteínas G, solo pueden activarse cuando unen Guanosin trifosfato (GTP). Por lo tanto la interacción del receptor unido al ligando provoca la activación de la proteína G y su unión al GTP. La proteína G activada, provoca la activación de una enzima amplificadora. Esta enzima convierte las moléculas precursoras ricas en fosfato en los segundos mensajeros. Por ejemplo, la enzima amplificadora adenilato ciclasa convierte el ATP en AMPc, mientras que la enzima amplificadora fosfolipasa C corta el fosfolípido de membrana 4,5-difosfato fosfatidil inositol (PIP2) en diacilglicerol (DAG) e inositol trifosfato (IP3). Como dijimos anteriormente la proteína G tiene actividad GTPasa (degrada el GTP), es decir que pasado un tiempo la misma proteína G se desactiva, terminando con la señal. En el estado inactivo la proteína G esta unida a GDP.
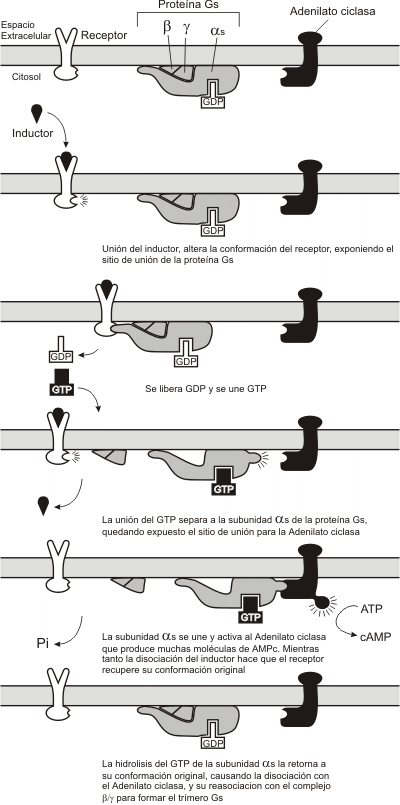
Fig. 7.5 - Secuencia de reacciones producidas a partir de la unión de la sustancia inductora con un receptor de membrana que activa a la proteína G, vía Adenilato ciclasa.
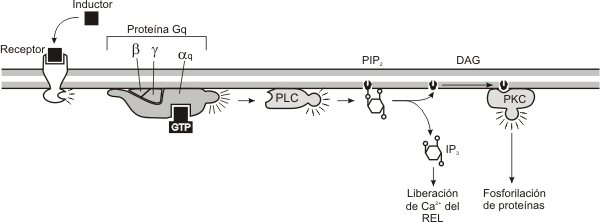
Fig. 7.6 - Secuencia de reacciones producidas a partir de la unión de la sustancia inductora con un receptor de membrana que activa a la proteína G, vía Fosfolipasa C (vía de los Fosfato inositoles).
|
Cuadro 7.1- Cuadro comparativo de las vías de transmisión a través de segundos mensajeros |
|||
|
Localización celular |
Vía Adenilato ciclasa (AC) |
Pasos generales |
Vía de los fosfato de Inositol |
|
Espacio extracelular |
Adrenalina ¯ |
Inductor (Primer mensajero) ¯ |
Adrenalina ¯ |
|
Membrana plasmática |
Receptor b-adrenérgico ¯ Proteína Gs ¯ Adenilato ciclasa (AC) ¯ |
Receptor ¯ Transductor ¯ Amplificador ¯ |
Receptor a1-adrenérgico ¯ Proteína Gq ¯ Fosfolipasa C (PLC) ¯ |
|
Citosol |
ATP ¯ AMPc ¯ Proteinquinasa A (PKA) ¯ Fosforilación de Fosforilquinasas ¯ Glucógeno ® Glucosa |
Precursor Fosforilado ¯ Segundo mensajero ¯ Fosforilación de Proteinquinasas ¯ Fosforilaciónes enzimáticas ¯ Respuesta Celular |
PIP2 ¯ DAG - IP3 - Ca2+ ¯ Proteinquinasa C (PKC) ¯ Liberación de Ca2+ al citosol ¯ Vasoconstricción |
Resumiendo, existen dos rutas principales de transmisión por medio de segundos mensajeros:
La primera vía utiliza como segundo mensajero al adenosin monofosfato cíclico (AMPc). El AMPc es generado por la enzima amplificadora Adenilato ciclasa.
La segunda vía utiliza una combinación de tres segundos mensajeros: iones calcio (Ca2+), inositol trifosfato (IP3) y diacilglicerol (DAG). En este caso la enzima amplificadora es la fosfolipasa C que genera el IP3 y el DAG a partir del fosfolípido de membrana el fosfatidil inositol difosfasto (PIP2). El IP3 provoca la liberación del Ca++ intracelular, de sus reservorios, como por ejemplo el REL.
Existen dos tipos de Proteínas G, las proteínas G estimuladoras (Gs y Gq) y las proteínas G inhibitorias (Gi)
La Proteína Gs (s, stimulatory G protein) unida a GTP activa a la AC (adenilato ciclasa) aumentando la cantidad de AMPc en el interior celular.
La proteína Gi (i, inhibitory G protein) unida a GTP inactiva a la adenilato ciclasa, disminuyendo indirectamente la cantidad de AMPc intracelular.
La proteína Gq unida a GTP activa a la fosfolipasa C, aumentando la cantidad de DAG, IP3 y Ca++ intracelular.
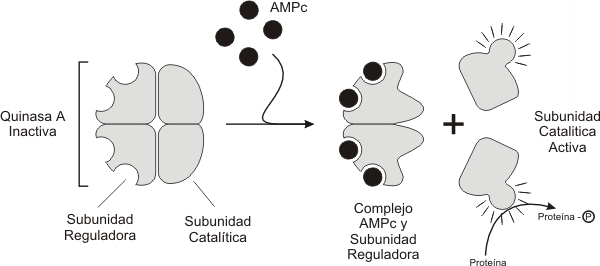
Fig. 7.7 - Activación de la proteinaquinasa A dependiente de AMPc
El AMPc regula la actividad de la proteinquinasa A (PKA)
Como vimos anteriormente la activación de la AC (adenilato ciclasa) por una proteína Gs aumenta la concentración de AMPc en el citosol. Este AMPc puede unirse a un sitio regulador de una proteinquinasa especifica denominada proteinquinasa A (PKA). Toda proteinquinasa A consta de dos subunidades una catalítica y otra regulatoria. La unión del AMPc a la subunidad regulatoria, provoca la activación de la PKA y la liberación de las subunidades catalíticas activas. Esta proteinquinasa inicia una cascada de fosforilaciones que determinan las respuestas celulares especificas de cada tipo celular, como se observa en el ejemplo de la Fig. 7.8.
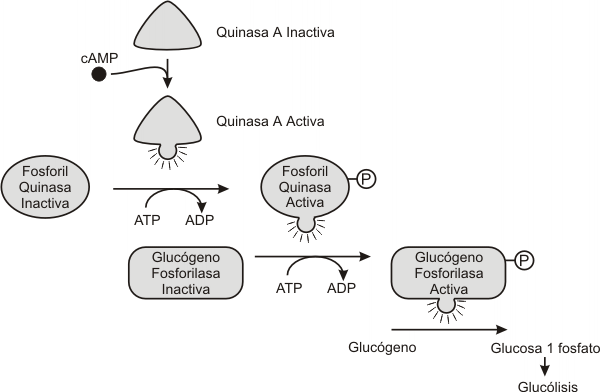
Fig. 7.8 - Efecto de la proteinquinasa A sobre la gluconeogénesis
EL diacilglicerol (DAG) activa a la proteinquinasa C (PKC)
La proteinquinasa C (por Ca2+ dependiente) es una enzima de membrana activada por el DAG. La PKC es una serin-treonin quinasa (agrega fósforo a los aminoácidos serina y treonina), que inicia una cadena de fosforilaciones, cuyos productos finales actúan a nivel del núcleo celular. Allí actúan como factores de transcripción celular que regulan la multiplicación celular. Cuando el DAG se degrada la PKC se inactiva.
El Inositol trifosfato (IP3), provoca la liberación de Ca2+ del retículo endoplásmico liso (REL)
EL IP3 provoca la apertura de los canales de Ca2+ dependientes de ligando (en este caso el IP3) del REL (retículo endoplásmico liso). Esto provoca la salida del Ca2+ del REL hacia el citosol. El calcio citosólico se comporta como segundo mensajero.
El Ca2+ citosólico se une a la calmodulina
La calmodulina es una proteína pequeña que une calcio. La unión del calcio a la calmodulina provoca un cambio conformacional en esta proteína. El complejo calcio-calmodulina se une a otras proteínas, activándolas. De esta forma el calcio por intermedio de su unión a la calmodulina puede actuar sobre varias vías de señalización. Por ejemplo, el complejo calcio-calmodulina puede unirse a una quinasa, calcio dependiente, para iniciar una cascada de fosforilaciones o a la enzima fosfodiesterasa que degrada el AMPc.
Ejemplos de respuestas inducidas por AMPc
Activación génica: La activación de la proteinquinasa A (PKA) por el AMPc, provoca la fosforilación de un factor de transcripción denominado, CREB (por elemento relacionado a proteínas que responden al AMPc) en las células que secretan el péptido somatostatina (hormona inhibidora de la hormona del crecimiento). El CREB fosforilado (CREBP) se une al ADN en sitios específicos denominados amplificadores regulados por AMPc, activando la transcripción de los genes que codifican esta hormona.
Sentido del olfato. Este sentido depende de receptores que responden a moléculas inductoras denominadas odorantes, que se encuentran en el aire. Los receptores de los odorantes de encuentran ubicados en neuronas ciliadas, que forman el epitelio olfatorio. Estas neuronas cuando mueren son reemplazadas regularmente por otras nuevas que se reproducen en el epitelio basal. El odorante se une al receptor, que es una proteína multipaso, y esto provoca la activación de una proteina G, asociada al receptor. Esto a su vez produce la activación de la enzima Adenilato ciclasa, con la consiguiente producción de AMPc (segundo mensajero) a partir del precursor fosforilado ATP. El aumento del AMPc en el citosol provoca la apertua de los canales de Na+ metabotrópicos. La apertura de estos canales permite la entrada de Na+ al interior celular, lo que provoca la despolarización de la membrana y la eventual generación de un potencial de acción. El potencial de acción es conducido por el nervio olfatorio hasta el cerebro, donde la señal es evaluada como un olor determinado.
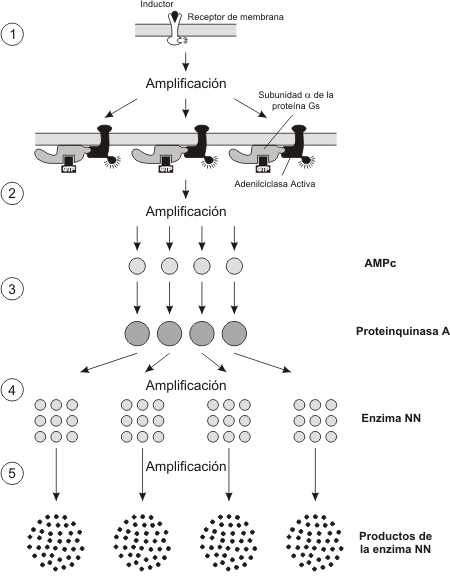
Amplificación de señales
La unión del inductor al receptor de membrana activa a varias proteínas G, cada proteína G puede activar a su vez una AC por un período prolongado, generándose muchas moléculas de AMPc, cada molécula de AMPc activa una proteinquinasa A, que a la vez pueden fosforilar muchas moléculas de enzima, activándolas. Cada enzima puede producir muchas moléculas de producto.
De esta simple secuencia deducimos, que de la unión de un inductor a su receptor de membrana, se obtiene una respuesta celular amplificada, pues obtenemos varias unidades de producto, partiendo de una unidad de inductor.
En algunos casos, la disociación entre el receptor y el ligando es tan rápida que no tiene lugar esta amplificación. En general las respuestas pueden ser rápidas, sólo si el mecanismo de inactivación también es rápido.
Inducciones en las que participan receptores de membrana con actividad enzimática
Los receptores de membrana con actividad enzimática, poseen en general tres dominios:
· Un dominio extracelular (extracitoplasmático), que une al primer mensajero (ligando)
· Un dominio transmembrana
· Un dominio intracelular (citoplasmático), con actividad enzimática.
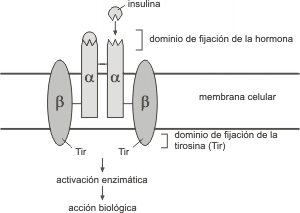
Fig. 7.10- Esquema de un receptor tirosinquinasa (RTK) de la insulina
Esta actividad enzimática es en general una quinasa.
En este caso nos referiremos a los receptores que cuando se activan por unión del ligando, la quinasa activada es una tirosinquinasa, es decir una enzima que fosforila específicamente aminoácidos tirosina. La actividad tirosinquinasa del receptor puede fosforilar tirosinas localizadas en el receptor (autofosforilación), como aminoácidos tirosina de otras proteínas citoplasmáticas.
La generación de múltiples señales simultaneas a partir de la activación de los receptores tirosinquinas (RTK), depende de tres factores:
· Organización Modular en la generación de señales. Los receptores activados fosforilan residuos de tirosina. Estos aminoácidos fosforilados son reconocidos por múltiples proteínas que poseen dominios SH2 (se unen a fosfotirosinas). Estas proteínas al unirse al receptor se activan y generan señales intracelulares.
· Moléculas Adaptadoras sin actividad enzimática, que se unen a los receptores por sus dominios SH2. Estas proteínas enganchan a su vez otras proteínas a los receptores activados. Estas proteínas unidas al receptor por medio de los adaptadores, activan nuevas vías de señalización.
· Proteínas Scaffolds (andamio, armazón, soporte) que permiten la activación simultanea (coordinada) de múltiples vías de señalización.
El receptor de insulina
Entre los RTK mas importantes encontramos al receptor de insulina. Recordemos que la insulina cumple múltiples funciones, es hipoglucemiante es decir que permite la entrada de glucosa a los tejidos insulinodependientes, disminuyendo de esta forma la cantidad de glucosa en sangre. Es un potente estimulante de la síntesis de lípidos en las células adiposas. También potencia la síntesis proteica y estimula el crecimiento y la división de todas las células del organismo.
Como vimos anteriormente el receptor de insulina se autofosforila en el aminoácido tirosina y fosforila también a otras proteínas que se asocian a él del lado citoplasmático. Estos sitios fosfotirosina sirven de enganche a proteínas que poseen dominios llamados SH2. La interacción de estas proteínas que poseen dominios SH2 y el receptor de insulina puede activar diferentes respuestas dependiendo de la proteína en particular. Si se trata de una molécula con actividad enzimática puede activarse, en cambio si se trata de una molécula adaptadora puede activar otras proteínas que se unen a ella.
La estructura del receptor de insulina es tetramérica. Dos subunidades alfa y dos subunidades beta. Las subunidades alfa unen la insulina y las subunidades beta, atraviesan la membrana y poseen la actividad tirosinquinasa.
Otros receptores con actividad tirosinquinasa
Entre otros RTKs podemos nombrar a los receptores del factor de crecimiento epidérmico (EGF) y el factor de crecimiento derivado de plaquetas (PDGF). Estos receptores a diferencia del receptor de insulina son monoméricos, mientras no están unidos al inductor. Cuando se activan, por unión del ligando, interactúan entre si para formar dímeros. La dimerización activa la función tirosinquinasa y la siguiente autofosforilación del receptor.
Proteina Ras
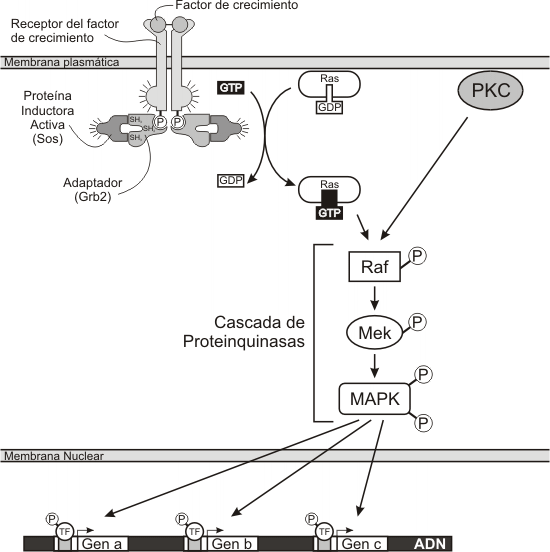
La proteína Ras es una pequeña proteína G citosólica. Es monomérica a diferencia de la proteína G de membrana que es trimérica. Al igual que otra proteínas G, tiene actividad GTPasa y por lo tanto muestra ciclos activos (unidos al GTP) e inactivos (unidos al GDP).
Esta proteína cumple un rol fundamental en varias vías de señalización internas. Una de las más importantes vías en la que interviene Ras es la cascada de proteinquinasa activada por mitógeno (MAPK). En esta vía un mitógeno (insulina, algún factor de crecimiento), activa a su RTK que se autofosforila, esto crea sitios fosfotirosina que actúan de anclaje para proteínas que poseen dominios SH2. En este caso se une al receptor, un complejo adaptador cuya función es activar a la proteína Ras. La proteína Ras activada (Ras-GTP), estimula a su vez a una tirosinquinasa llamada Raf que inicia una cadena de fosforilaciones, que culmina con la activación de genes que están involucrados en la síntesis de ADN y en la activación de la división celular.
Inducciones en las que participan receptores citosólicos
Las hormonas esteroideas, tiroxina (T4) y triiodotironina (T3) , calcitriol (vitamina D) y el ácido retinoico son ejemplos de inductores que tienen sus receptores en el citosol de las células inducidas. Los tres primeros se vehiculizan por la sangre y entran en la categoría de inductores endocrinos, mientras que el ácido retinoico interviene en inducciones parácrinas, sobre todo durante el desarrollo embrionario. En el citosol, el inductor se une a su correspondiente receptor, formando un complejo que ingresa en núcleo uniéndose a la secuencia reguladora de un gen específico, conocida como elemento de respuesta a la hormona, el cual se activará, desencadenándose la transcripción del mismo. Como resultado se formará un ARNm y a partir de este la síntesis de una proteína, como respuesta de la célula inducida.
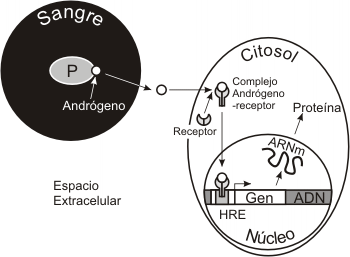
Fig. 7.12 - Inducción celular a través de un receptor citosólico. Modo de acción de las hormonas esteroides, T3 y T4, calcitrioll y ácido retinoico.
El óxido nitrico (NO) como inductor
Otro ejemplo, lo constituye el oxido nítrico (NO). Este último cuando es secretado por las células endoteliales de los vasos sanguíneos o por algunas neuronas, se comporta como un inductor. Su acción dentro de la células es muy breve, pues es metabolizado en el lapso de breves segundos.
El óxido nítrico secretado por las células endoteliales tiene como blanco a las células musculares lisas de los mismos vasos, las cuales se relajan, produciendo por lo tanto una vasodilatación.
Durante el proceso de erección del pene, la acetilcolina es liberada por los terminales axónicos del sistema parasimpático e interactúa con los receptores de membrana de las células endoteliales. Como respuesta se activa en estas células la enzima óxido nítrico sintetasa que genera óxido nítrico a partir del aminoácido arginina, este inductor pasa al espacio intercelular hasta alcanzar el citoplasma de las células musculares lisas, promoviendo la vasodilatación y la consiguiente erección del pene.
Otro ejemplo es el de la nitroglicerina, utilizada para tratar la angina de pecho, una afección cardiaca. Luego de su administración la nitroglicerina se convierte gradual y lentamente en óxido nítrico, que dilata los vasos coronarios por períodos relativamente largos.
Un descubrimiento reciente, es la participación del oxido nítrico, en el proceso de fertilizacion. En este complejo proceso el citoplasma del espermatozoide posee la enzima oxido nítrico sintetasa (NOS), que se activa con la reacción acrosómica, de esta forma se activa la síntesis del NO. Una vez producida la fusión entre el óvulo y el espermatozoide, tanto la enzima que lo sintetiza como el NO son liberados dentro de la célula huevo, donde el NO produce la liberación del Ca2+ intracelular en el citoplasma, acontecimiento que activa al zigoto que comienza a dividirse y crecer en un embrión.
Actividades de AutoevaluaciÓn
1) ¿Qué es y en qué consiste la transducción de una señal?
2) ¿Cuáles son los pasos desde la síntesis del AMPc en la monocapa citosólica de la membrana plasmática del hepatocito hasta la liberación de la glucosa al torrente sanguíneo?
3) ¿Qué es la amplificación de una señal? ¿Cuál es la diferencia con la transducción de una señal?
4) ¿De que manera la reacción en cascada produce la amplificación de la señal? ¿Cómo incrementa las posibilidades de regulación metabólica?
5) ¿Qué determina si un estímulo que actúa a través de una proteína G será estimulador o inhibidor para un efector?
6) ¿Qué estímulos extracelulares conducen a la formación de I3P? ¿Cuál es el mecanismo de formación de este segundo mensajero?
7) Cuál es la relación entre la formación de I3P y el aumento del CA++ intracelular?
8) ¿Cómo altera la cascada del AMPc la traducción y la transcripción una célula?
Preguntas multiple opción1) La función del AMPc es la de:
a- primer mensajero
b- segundo mensajero
c- transportador de electrones
d- transportador de energía
2) Las hormonas esteroides tienen sus receptores en:
a- la monocapa intracelular de la membrana plasmática
b- la monocapa intracelular de la membrana plasmática
c- el citoplasma
d- las chaperonas
3) ¿Cuál de las siguientes hormonas disminuye la concentración de ázucar en sangre?
a- Glucagón
b- Aldosterona
c- Insulina
d- Todas las hormonas esteroideas
4) ¿Cuál de las siguientes puede representa la secuencia precisa de componentes en una respuesta celular a una hormona peptídica?
a-
Hormona
unida a la adeniciclasa (AC) ![]() proteína G
proteína G ![]() proteinquinasa
proteinquinasa ![]() fosforilación de enzimas
fosforilación de enzimas
b-
Hormona
unida al receptor ![]() proteína G
proteína G ![]() factor de transcripción
factor de transcripción![]() proteinquinasa
proteinquinasa
c-
Hormona
unida a proteina G ![]() AC
AC ![]() proteinquinasa
proteinquinasa ![]() fosforilación de proteínas
fosforilación de proteínas
d-
Hormona
unida al receptor ![]() proteína G
proteína G ![]() AC
AC ![]() proteinquinasa
proteinquinasa ![]() fosforilación de proteínas
fosforilación de proteínas
5) Un segundo mensajero derivado de la estructura lipídica de la membrana plasmática es:
a- AMPc
b- Calmodulina
c- IP3
d- Ca++
6) La principal diferencia en el mecanismo de acción entre las hormonas esteroideas y peptídicas es que:
a- Las hormonas esteroideas principalmente afectan la síntesis proteica mientras que las peptídicas afectan mayormente la actividad de las proteínas ya existentes en la célula
b- Las células blanco reaccionan más rápido a las hormonas esteroideas que peptídicas
c- Las hormonas esteroideas entran en el núcleo mientras que las peptídicas permanecen en el citoplasma
d- Las hormonas esteroideas se unen a un receptor proteico mientras que las peptídicas se unen a la proteína G.
Bibliografía
· Alberts, B. et al; (1996) Biología Molecular de la Célula; 3° Edición; Ediciones Omega S.A. Barcelona.
· Karp, G.; (1998) Biología Celular y Molecular; Ed. Mc Graw Hill Interamericana. México.
· De Robertis (h), Hib, J.; Ponzio, R.; (1996) Biología Celular y Molecular de De Robertis; 12° Edición; El Ateneo. Bs.As.
· De Robertis, E; Hib, J.; (1998) Fundamentos de Biología Celular y Molecular; El Ateneo. Bs.As.
· Linder M. and Gilman A. (1992). G proteins. Sci. Am. 267 (1): 56-61.
· Scott, J. And Pawson, T. (2000). Cell communication: The Inside Story. Sci. Am. 282 (6): 72-79
· Smith and Wood; (1997) Biología Celular; Ed. Addison-Wesley, Iberoamericana S.A.
[1] Se denominan sinapsis a las uniones donde el axón o alguna otra porción de una célula (la célula presinática) termina en las dendritas, soma o axón de otra neurona o en una célula muscular o glandular (célula posináptica)